










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkFave. Sección ciencias agrarias
versão impressa ISSN 1666-7719
FAVE. Secc. Cienc. agrar. vol.14 no.2 Santa Fe dez. 2015
ARTICULO ORIGINAL
Generación de biomasa y rendimiento en trigo frente a infecciones de Puccinia triticina y Pyrenophora tritici-repentis
SCHIERENBECK, M; FLEITAS, M.C.; GOLIK, S. I. & SIMÓN M.R.
1.- Cátedra de Cerealicultura. Facultad de Ciencias Agrarias y Forestales, UNLP. Av. 60 y 119 s/n. La Plata,
Buenos Aires, Argentina (CP: 1900). Tel.: +54 (0221) 423-6758. Email: Email: m_schierenbeck@hotmail.com
2.- CONICET CCT La Plata. Calle 8 Nº 1467. La Plata, Buenos Aires, Argentina. Tel.: +54 (0221) 457-6423.
Manuscrito recibido el 27 de julio de 2015 y aceptado para su publicación el 11 de noviembre de 2015.
RESUMEN
El objetivo del presente trabajo fue evaluar el efecto de un patógeno fúngico biotrófico (Puccinia triticina), causal de la roya de la hoja (RH) y un necrotrófico (Pyrenophora tritici-repentis), causal de la mancha amarilla (MA) sobre los atributos fisiológicos vinculados a la generación de biomasa y rendimiento en trigo. Se condujo un ensayo durante 2012 y 2013 utilizando un diseño experimental en parcela sub-dividida con tres repeticiones. Las parcelas principales fueron las inoculaciones con los patógenos. Las sub-parcelas fueron: 1-Sin inóculo y aplicación de fungicida, 2-baja concentración de inóculo y 3-alta concentración de inóculo. Como sub-sub-parcela se sembraron 10 cultivares de trigo. Los resultados indican que MAafectó más la absorción de radiación a partir de mayores niveles de senescencia foliar, en tanto que RHafectó más la eficiencia en el uso de la radiación, probablemente por la interacción particular que los patógenos biotróficos presentan con las células del hospedante.
Palabras claves: Radiación absorbida; Eficiencia de uso de la radiación; Patógenos foliares; Trigo
SUMMARY
Wheat biomass and yield generation under Puccinia triticina and Pyrenophora tritici-repentis infections
The present study was designed to determine the effects of inoculations of foliar disease pathogens with different nutritional habits: Pyrenophora tritici-repentis (necrothroph -PTR-) and Puccinia triticina (biotroph -LR-) on the physiological components of biomass generation, like absorbed photosynthetic active radiation (APAR) and radiation use efficiency (RUE), and on the yield production in wheat. Field experiments were carried out in 2012-2013 at FCAyF-UNLP using a split-split-plot design with three replications. The main plots were the pathogen inoculations. Subplots were inoculation treatments: 1- Without inoculation, 2- Low dose of spores concentration inoculation, 3- High dose of spores concentration. Ten cultivars were the sub-subplots. Foliar diseases decreased biomass production mainly explained by reductions in green leaf area, healthy area duration and reductions on APAR, showing higher declines under PTR inoculations. A detrimental effect in the RUE was detected at LR inoculations, which could be associated with the nutritional habit of the pathogen.
Key words: Biomass; Radiation use efficiency; Radiation absorption; Foliar diseases; Wheat
INTRODUCCIÓN
Las enfermedades foliares constituyen uno de los principales factores bióticos que limitan la expresión de los rendimientos y calidad del cultivo de trigo en todo el mundo (11). En Argentina, se considera que las de mayor importancia relativa son aquellas enfermedades fúngicas que afectan tejidos foliares como la roya de la hoja (agente causal Puccinia triticina Eriks) y la mancha amarilla [Pyrenophora tritici-repentis (Died.) Drechs., anamorfo Drechslera tritici-repentis) (Died.) Shoem. ] La mancha amarilla (MA) es causada por un patógeno necrotrófico por lo que obtiene los nutrientes necesarios para su crecimiento de tejido muertos, luego de matar al hospedante por secreción de toxinas que generan la degradación de las paredes celulares y produciendo consecuentemente las características lesiones cloróticas y necróticas (10). Por otro lado, la roya de la hoja (RH) es causada por un patógeno biotrófico que obtiene los nutrientes exclusivamente de tejidos vivos y utiliza estos recursos para su crecimiento y esporulación (19), caracterizándose por producir profundos cambios en la fisiología de los hospedantes, ya que alteran el metabolismo de las plantas al parasitar las células vivas.
La producción de biomasa para un amplio rango de cultivos como el trigo (Triticum aestivum L.) depende de la capacidad del canopeo de (i) interceptar la radiación incidente (eficiencia de intercepción, Ei), que es función del índice de área foliar (IAF) y la arquitectura del canopeo (coeficiente de extinción k) y (ii) la eficiencia con que se convierte la energía lumínica en biomasa (eficiencia en el uso de la radiación, EUR). Diversos autores afirman que las enfermedades afectan principalmente la Ei, principalmente por reducciones del área verde y aumentos de la intercepción de radiación por área foliar enferma que no podrá ser utilizada para la generación de biomasa (5). El efecto de las enfermedades sobre la EUR presenta resultados contradictorios en la bibliografía: sin embargo, algunos trabajos indican que enfermedades originadas por patógenos biotróficos podrían disminuir la EUR (13).
El grado de daño que producirá una determinada enfermedad dependerá no sólo de la incidencia y/o severidad del patógeno, sino también del impacto sobre los atributos del cultivo responsables de la asimilación de carbono en el sistema (20). En ese sentido, algunos autores sostienen que el estudio del impacto de los patógenos sobre variables relacionadas a la ecofisiología del cultivo como el IAF verde (IAFV), duración del área foliar verde (DAFV) y sobre la proporción de IAF no verde (IAFNV) en relación al IAF total (%AFNV) (16) en varios estadios permitiría disminuir la variabilidad e incrementar la predectibilidad y extrapolaridad en las estimaciones de pérdida de biomasa y rendimiento (4). Hasta el presente, no se dispone de antecedentes a nivel mundial que evalúen el impacto de P.triticina(biotrófico) y P.tritici-repentis(necrotrófico) inoculados en forma separada en un mismo ambiente sobre los atributos ecofisiológicos vinculados a la generación de biomasa y rendimiento. En base al efecto diferencial que patógenos biotróficos y necrotróficos pueden ejercer con las células del hospedante se hipotetiza que los primeros podrían tener mayor incidencia sobre la EUR, en tanto que los necrotróficos podrían afectar en mayor medida la absorción de radiación por parte del cultivo.
MATERIALES Y MÉTODOS
Datos climáticos y diseño del ensayo
Los ensayos se llevaron a cabo durante 2012 y 2013 en la E.E J. Hirschhorn, Facultad de Ciencias Agrarias y Forestales, Universidad Nacional de La Plata (34° 52' LS; 57° 58' LO). Durante el ciclo del cultivo se registraron diariamente las variables climáticas en la estación meteorológica situada a 200 m de los ensayos. El suelo fue un Argiudol típico con los siguientes resultados de análisis: 0-20 cm: Materia orgánica 3,3%; N 0,167%; Nitratos 26,3 ppm; P 16,1 ppm, pH 6,15. 20-40cm: Nitratos 22,5 ppm, pH 5,89
Se utilizó un diseño experimental en parcela sub-dividida con tres repeticiones. Las parcelas principales fueron las inoculaciones con: 1-Py.tritici-repentis y 2- P.triticina. La sub-parcela correspondió a los tratamientos de inoculación: 1-sin inóculo y aplicación de fungicida (SI), 2-con baja concentración de inóculo (BCI) y 3-con alta concentración de inóculo (ACI). Como sub-sub-parcela se sembraron 10 cultivares comerciales de trigo de ciclo intermedio-largo. La siembra se realizó a mediados del mes de junio con una sembradora experimental a 20 cm entre líneas y una densidad de 250 pl.m-2. Cada parcela experimental fue de 7,7 m2 (5,5 m x 1,4 m). Se aplicaron 100 kg/ha de N dividido en dos momentos (siembra y fines de macollaje) y 50 kg/ha de P a la siembra. Entre las sub-parcelas se realizó la intersiembra de Avena sativa para disminuir el traspaso de inóculo. En la parcela principal con P.triticina se sembró una bordura con una mezcla de cultivares susceptibles (Baguette 21 y Baguette 30) para facilitar la inoculación de las parcelas. Las parcelas principales estuvieron distanciadas al menos 200 m para minimizar el traspaso de inóculo.
Para la preparación del inóculo de Py. tritici-repentis se utilizó una mezcla de aislados obtenidos de diferentes regiones del país y se multiplicó mediante sucesivos repiques en flujo laminar en caldo V8R (agar-agar, caldo de verduras, carbonato de calcio). Mediante raspado de las cajas con agua destilada se extrajo el micelio infectivo y se realizaron conteos con cámara de Neubauer para ajustar una solución BCI (3x102 esporas/ml) y otra ACI (3x103 esporas/ml). Las inoculaciones se realizaron a fines de macollaje (EC24) (21) y en hoja bandera desplegada (EC39). Para ambas concentraciones se utilizaron 800 ml de solución (inóculo, agua destilada estéril y coadyuvante) por parcela y por aplicación. Como fuente de inóculo inicial de P.triticina se utilizó una mezcla de razas virulentas provistas por el Ing. Pablo Campos del INTA Bordenave. Las inoculaciónes se realizaron en EC24 y EC39 y consistieron en la pulverización de las borduras cercanas al tratamiento de ACI con 0,5 mg por planta de esporas y con 0,2 mg de esporas en BCI. En las sub-parcelas SI se aplicó el fungicida OrquestaTM Ultra (fluxapyroxad 50g/l, epoxiconazole 50 g/l y pyraclostrobin 81 g/l) en pleno macollaje (EC23) y en EC39 mediante una mochila de gas carbónico con pastillas de abanico plano. La tasa de aplicación fue de 140 l/ha con una dosis de 1,2 L/ha de producto.
Evaluaciones y análisis estadístico
Las evaluaciones se realizaron en EC39, antesis (EC60) y grano pastoso (EC82) y consistieron en la determinación de la severidad en todas las hojas con al menos una porción de tejido verde (% de lesión cubierta por la enfermedad en la hoja) en siete a diez plantas tomadas al azar por cada parcela. Se evaluó la biomasa, mediante extracción de 1,50 m lineales de plantas y posterior secado a 60°C por 72 hs de una alícuota para determinar la materia seca aérea. El índice de área foliar total se estimó mediante el conteo de macollos en 2 m lineales y la medición de todas las hojas con al menos una porción de tejido verde de un total de siete macollos y empleando un factor de forma de 0,835 (9). Mediante las estimaciones de severidad se calculó el IAFV y el % AFNV en las tres evaluaciones. Con las medias de IAFV de las tres evaluaciones se calculó la DAFV (20). La radiación incidente (RI) y la radiación fotosintéticamente activa interceptada (RFAi) fueron evaluadas mediante un ceptómetro y utilizadas junto a las evaluaciones de severidad para calcular la radiación absorbida (RFAa) por tejido verde. La eficiencia de uso de la radiación (EUR) se calculó como la pendiente de la regresión entre la RFAi o RFAa y la biomasa generada acumulada a lo largo del ciclo del cultivo. Se determinaron a cosecha los componentes de rendimiento: espigas.m-2 (NESP) mediante el conteo de 3m lineales en cada parcela y los granos.espiga-1 (NGE) trillando 20 espigas por parcela. Ambos componentes fueron utilizados para calcular el número de granos.m-2 (NGT). Por su parte, el peso de mil granos (PMG) se calculó mediante el conteo y peso de los granos obtenidos por la trilla de 20 espigas. Asimismo se cosecharon 5,5 m (1,1m2) como estimador del rendimiento a cosecha. El análisis de datos se realizó mediante un análisis de varianza (ANVA) para parcelas divididas mediante el programa GenStat 12th Edition. Las medias se compararon mediante el test LSD (P=0,05).
RESULTADOS
Porcentaje deÁrea foliar no verde (%AFNV), dinámica del índice de área foliar verde (IAFV) y duración de área foliar verde (DAFV)
Las variaciones climáticas entre años influenciaron fuertemente el desarrollo epidemiológico de las enfermedades evaluadas. En este sentido, el año 2012 presentó condiciones más favorables para el desarrollo de enfermedades en post-floración debido a la mayor acumulación de precipitaciones, humedad relativa y temperaturas medias más elevadas que provocaron mayores niveles de severidad en EC82, ABCPE, en el %AFNV (Tabla 1).
Tabla 1. Temperatura media, Precipitaciones, Humedad relativa y Radiación Incidente registrados durante el ensayo en la Estación Experimental J.Hirschorn (La Plata, Argentina).
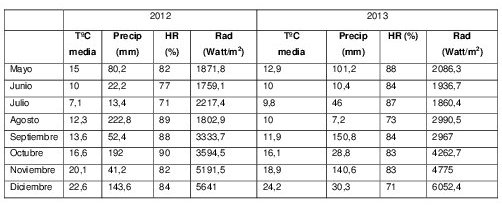
T°C media: Temperatura media; Precip: Precipitaciones; Rad: Radiación incidente; HR: Humedad relativa ambiente
En el estadio de EC39, el %AFNV mostró diferencias significativas para la interacción Patógeno x Inoculación (P=0,05). En este sentido, inoculaciones con Py.tritici-repentis presentaron un aumento significativo en esta variable en los tratamientos BCI y ACI. En contraposición, no se detectaron diferencias significativas cuando P.triticina fue inoculado. En la evaluación de EC60, el %AFNV presentó un aumento desde 16,7 % (SI) al 25,8 % (BCI) y 27,1 % (ACI) ante inoculaciones con Py.tritici-repentis y desde 24,3 % (SI) a 32,4 % (BCI) y 32,3 % (ACI) ante P.triticina. En EC82, los tratamientos BCI y ACI presentaron los mayores valores sin diferencias significativas entre ellos (79 y 73,6 %, respectivamente) frente a 53% (SI) cuando Py.tritici-repentis fue el patógeno principal. Para P.triticina, no se detectaron diferencias significativas entre los tratamientos BCI y ACI (79 y 78,5%, respectivamente), mientras que el tratamiento SI presentó los menores valores (62,4%) (Figura 1).

Figura 1. Porcentaje de área foliar no verde (%AFNV) promedio en los estadios EC39, EC60 y EC82 para la interacción Patógeno x Inoculación. Medias seguidas por la misma letra dentro de cada patógeno y estadio son significativamente iguales (LSD p=0,05)
El valor de IAFV de las tres evaluaciones mostró reducciones notorias con el aumento en la concentración de inóculo frente al tratamiento protegido (SI) (P<0,001) en promedio de los dos patógenos. En EC39 y con respecto al testigo SI (IAFV=7,05), los tratamientos BCI y ACI disminuyeron el valor de IAFV un 21,0 % y un 26,6 % respectivamente. En antesis, se detectaron disminuciones de 24,3 % (BCI) y 38,5 % (ACI) con respecto al tratamiento SI (IAFV=4,97). En tanto que para la evaluación de EC82 los porcentajes se disminución fueron de 51,6% (BCI) y 66,3% (ACI) frente al tratamiento SI (IAFV=0,694).
En la evaluación de EC60, el IAFV mostró diferencias significativas para la interacción Año x Patógeno x Inoculación (P=0,041). En los dos años evaluados, el tratamiento ACI fue el que presentó menores valores (Figura 2a). En 2012 y cuando Py.tritici-repentis fue el patógeno principal, los tratamiento BCI y ACI disminuyeron el valor de IAFV un 23,9 y 37,8 % con respecto al tratamiento SI (IAFV=4,76), respectivamente. En el mismo año y para P.triticina, se detectaron disminuciones de 24,2 (BCI) y 35,2 % (ACI) con respecto al tratamiento SI (IAFV=5,37). En 2013, los porcentajes de disminución fueron de 25,1 (BCI) y 44,0 % (ACI) frente al tratamiento SI (IAFV=5,57) en Py.tritici-repentis y del 23,4 (BCI) y 36,3 % (ACI) para con respecto al tratamiento SI (IAFV=4,19) de P.triticina (Figura 2a). En EC82 también se encontraron diferencias significativas para esta interacción (P=0,06). Tanto en 2012 como en 2013, el tratamiento SI presentó valores superiores de IAFV diferenciándose significativamente de BCI y ACI. El rango de valores de IAFV en 2012 fue para MA de 0,42 (SI) a 0,09 (ACI) implicando una disminución del 78,6% ante el tratamiento ACI. Para RH y teniendo en cuenta el mismo año, los valores fueron de 0,34 (SI) a 0,08 (ACI), involucrando una caída del 76,5 % en el IAFV (Figura 2b). En el año 2013, se presentaron valores superiores de IAFV para ambas enfermedades. Para Py.tritici-repentis, la disminución en el IAFV debido a mayores niveles de enfermedad variaron de 1,25 (SI) a 0,44 (ACI), implicando una caída del 64,8 %. Para P.triticina, los valores fueron desde 0,75 (SI) a 0,31 (ACI), correspondiente a una disminución en el IAFV del 58,7% (Figura 2b)
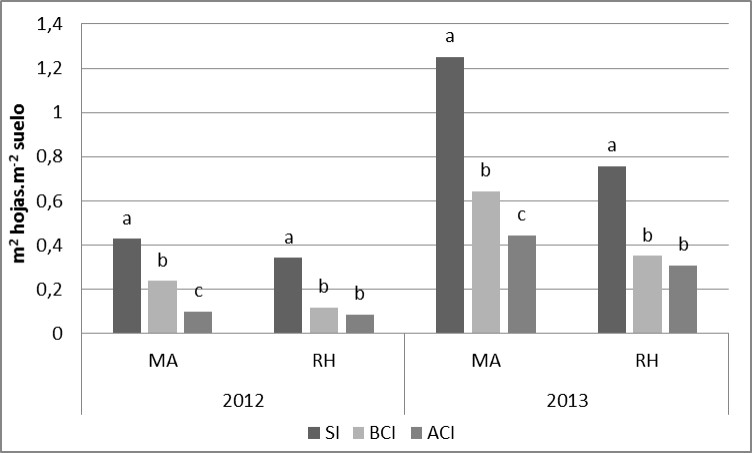
Figura 2a y 2b. Índice de área foliar verde en el estadio EC60 y EC82 para la interacción Año x Patógeno x Inoculación. Medias seguidas por la misma letra dentro de cada año y patógeno son significativamente iguales (LSD p=0,05)
Se detectaron diferencias significativas para la DAFV en la interacción Año x Patógeno x Inoculación (P=0,06). En ambos años, el tratamiento SI presentó valores superiores de DAFV diferenciándose significativamente de los restantes. El rango de valores en 2012 fue para MA de 189 (SI) a 121 días (ACI) implicando una disminución del 36,2% en la DAFV. Para RH y comparando los mismos tratamientos, la caída en el DAFV fue del 32,4% (214 vs. 145 días). En el año 2013 con inoculaciones con Py.tritici-repentis, la disminución en la DAFV debido a mayores niveles de enfermedad fue del 41,1% cuando se comparó el tratamiento ACI con el que SI (205 días). Para P.triticina, la caída en el DAFV fue del 31,9 % cuando se comparó el tratamiento ACI con el SI (167 días) (datos no mostrados).
Influencia de la mancha amarilla y la roya de la hoja sobre la generación de biomasa y sus atributos
La intercepción (RFAi) y absorción de radiación (RFAa) evidenciaron diferencias significativas para la interacción Año x Inoculación (Tabla 2)revelando que incrementos en la concentración de inóculo de los patógenos provocaron disminuciones en ambas variables. La RFAi evidenció en 2012 una caída del 4,68 % (ACI) con respecto al tratamiento SI, en tanto que en 2013 las disminuciones fueron mayores: 4,61 (BCI) y 10,9 % (ACI). Por su parte, la RFAa mostró reducciones con respecto al tratamiento SI del 6,33 (BCI) y 14,4 % (ACI) en 2012 y del 13,8 (BCI) y 18,3 % (ACI) con respecto al tratamiento SI en 2013 (Figura 3). La interacción Patógeno x
Inoculación para la RFAi y RFAa en EC1-EC82 (Tabla 2) manifestó mayores caídas porcentuales ante MA frente a aumentos en la concentración de inóculo. La RFAa presentó reducciones de hasta 20,1 % (ACI) con respecto al tratamiento protegido (754 MJ.m-2) ante inoculaciones de Py.tritici-repentis en comparación a P.triticina, donde las reducciones llegaron al 12,7 % (ACI) con respecto a SI (682 MJ.m-2) (datos no mostrados).
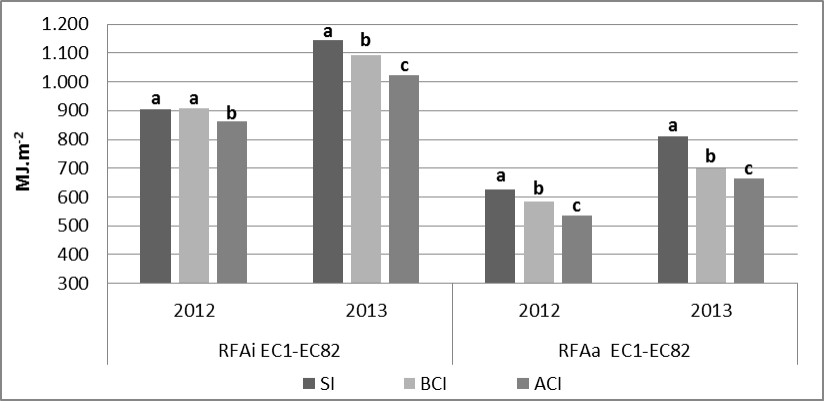
Figura 3. Radiación fotosintéticamente activa interceptada y absorbida promedio en el periodo EC1-EC82 para la interacción Año x Inoculación. Medias seguidas por la misma letra dentro de cada año son significativamente iguales (LSD p=0,05).
Tabla 2. Valores p de los cuadrados medios del ANVA de Radiación fotosintéticamente activa interceptada (RFAi) y absorbida (RFAa), eficiencia en el uso de la radiación interceptada (EURint) y absorbida (EURabs) y biomasa aérea en los estadios de hoja bandera desplegada (EC39), Antesis (EC60) y grano pastoso (EC82)

En cuanto a la producción de biomasa, en EC39 y comparado con el tratamiento SI, se evidenció una reducción del 11,5 % ante el tratamiento ACI. Para EC60, se detectaron disminuciones en la producción biomasa de 7,8 (BCI) y 19,2 % (ACI) con respecto al tratamiento SI. En tanto que para la evaluación de EC82 los porcentajes de disminución fueron de 7,9 (BCI) y 20,2% (ACI) frente al tratamiento SI (Figura 4). Se detectaron menores niveles de producción de biomasa para Py.tritici-repentiscon respecto a P.triticina en EC39 (540 g.m-2 vs. 715 g.m-2) y EC60 (1037 g.m-2 vs. 1238 g.m-2), en tanto que en EC82, P.triticina mostró los menores valores (1501 g.m-2 vs. 1374 g.m-2).
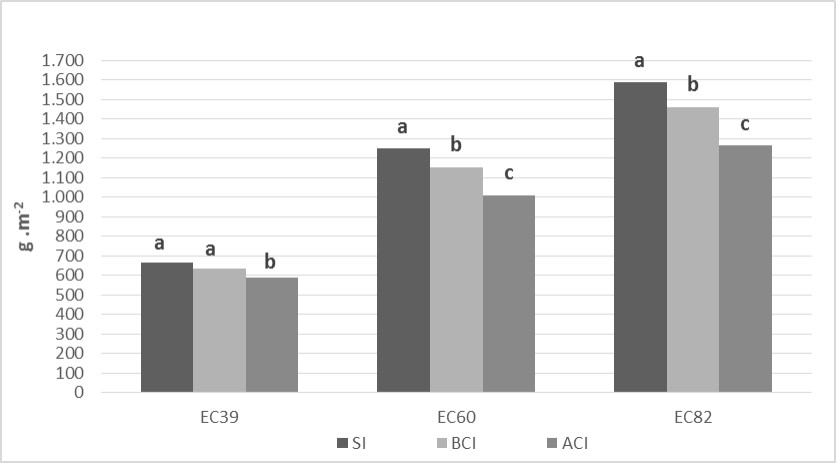
Figura 4. Biomasa aérea promedio en los estadios EC 39, EC 60 y EC 82 para el factor Inoculación. Medias seguidas por la misma letra dentro de cada estadío son significativamente iguales (LSD p=0,05)
Con respecto a la EURabs y EURint en la interacción Patógeno × Inoculación se observó una tendencia a mayores reducciones porcentuales ante incrementos en la concentración de inóculo cuando P.triticina fue el patógeno inoculado. En ese sentido, la EURabs no presentó disminuciones con el incremento en la concentración de inóculo de Py. tritici-repentis, en tanto que con P. triticina disminuyó 11,2 %. Por su parte, en la EURint se observó una mayor reducción en RH (23 %) que en MA (15,6 %) en MA cuando se comparó el tratamiento ACI con el SI (Figura 5).
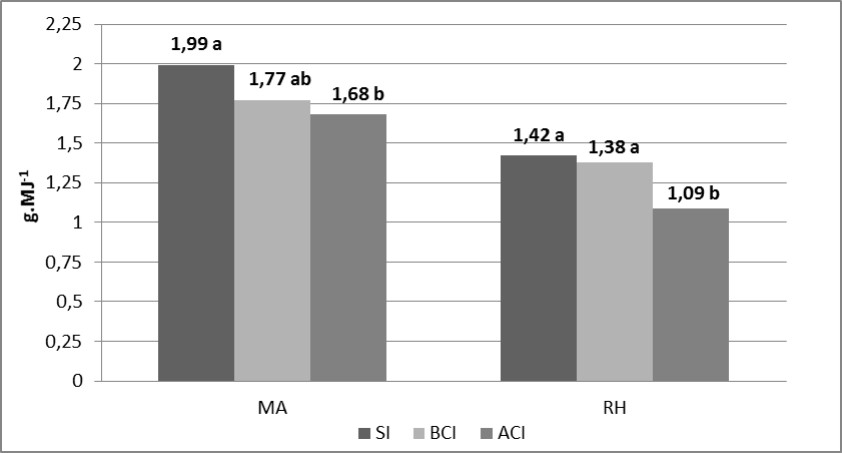
Figura 5. Eficiencia en el uso de la radiación interceptada para la interacción Patógeno x Inoculación. Medias seguidas por la misma letra dentro de cada patógeno son significativamente iguales (LSD p=0,05)
Se hallaron diferencias significativas para la interacción Año x Patógeno para la variable EURint (Tabla 2). En 2012, inoculaciones con P.triticina generaron caídas en esta variable con respecto a Py.tritici repentis de 35,6 % (1,22 g.MJ-1 vs. 1,90 g.MJ-1), por su parte, en 2013, estas disminuciones fueron del 20,8 % (1,38 g.MJ-1 vs. 1,74 g.MJ-1). En el mismo sentido, la EURabs mostró una tendencia a mayores reducciones de esta variable frente a inoculaciones con P.triticina en relación a Py.tritici-repentis en ambos años. Estas disminuciones llegaron hasta 24,9 % en 2012 (2,51 g.MJ-1 vs. 3,34 g.MJ-1) y en torno al 13,2 % en 2013 (2,53 g.MJ-1 vs. 2,92 g.MJ-1).
Rendimiento y sus componentes
Se presentaron diferencias significativas para la interacción Año x Inoculación para el NGE (Tabla 3). En 2012 y comparado con el tratamiento SI (35,9 granos.espiga-1), los tratamientos BCI y ACI disminuyeron el NGE un 6,65 y 8,33 %, respectivamente. En 2013 se detectaron caídas más pronunciadas en esta variable ante aumentos en la concentración de inóculo con disminuciones de 17,4 (BCI) y 22,4 % (ACI) con respecto al tratamiento SI (40 granos.espiga-1). El PMG exhibió diferencias significativas para la interacción Patógeno x Inoculación (Tabla 3). Cuando Py. tritici-repentis fue el patógeno principal, se detectó un merma en el PMG frente al tratamiento SI (34,4 g) de 6,25 (BCI) y 4,30 % (ACI). Para inoculaciones de P.triticina, la caída del PMG frente a aumentos en el nivel de inóculo fueron mayores: 9,40 (BCI) y 8,07 % (ACI) con respecto a SI (36,2 g). Los tratamientos de Inoculación afectaron significativamente el NGT y el rendimiento (Tabla 3). Cuando se comparó el tratamiento SI con el de ACI, el NGT disminuyó de 17582 granos.m-2 a 15044 granos.m-2, implicando una merma del 14,4 %. El rendimiento evidenció reducciones significativas ante los tratamientos BCI y ACI, sin presentar diferencias significativas entre éstos. Con respecto al tratamiento protegido SI (5654 kg.ha-1), la reducción en el rendimiento fue del 17,7 % para el tratamiento BCI y del 23,8 % con respecto al tratamiento ACI (Figura 6).
Tabla 3. Valores p de los cuadrados medios del ANVA de rendimiento y componentes del rendimiento
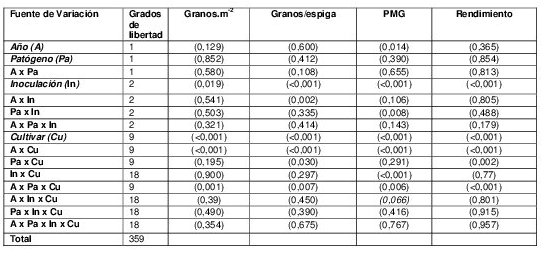
Granos.m-2= Granos por metro cuadrado; PMG: Peso de mil granos

Figura 6. Rendimiento promedio para el factor Inoculación. Medias seguidas por la misma letra son significativamente iguales (LSD p=0,05)
DISCUSIÓN
Las enfermedades foliares redujeron la biomasa aérea con respecto a tratamientos protegidos, al afectar el IAFT y IAFV a causa de senescencia foliar anticipada, disminuyendo consecuentemente la RFAi, concordando con lo reportado por diversos autores (1,14,16). A su vez, parte de la radiación interceptada fue capturada por área foliar enferma o senescente (IAFNV), produciéndose una reducción en la radiación que es efectivamente absorbida por área foliar verde y funcional (RFAa) y que posteriormente es utilizada para producir fotoasimilados para la generación de biomasa y rendimiento. Bajo las condiciones evaluadas en este estudio, incrementos en la concentración de inóculo de Py.tritici-repentis provocaron reducciones significativamente superiores tanto en el IAFT-IAFV, como en la RFai y RFAa con respecto a P.triticina.
Sumado a esto, la EUR evidenció un efecto diferencial dependiente del hábito nutricional del patógeno inoculado. En este sentido, se detectaron mayores reducciones en la EURint y EURabs ante inoculaciones del patógeno biotrófico P.triticina que coincide con lo reportado por otros autores (15). Si bien varios trabajos afirman que la EUR no se ve modificada por las enfermedades foliares al ser un atributo más conservativo (1, 20), diversos autores afirman que el hábito nutricional de los patógenos fúngicos podría ejercer efectos diferenciales sobre este atributo (2). En términos generales, se señala que infecciones causadas por patógenos necrotróficos como Py.tritici-repentis no generan reducciones en la EUR al no interactuar con células vivas del hospedante, ya que el patógeno las degrada previamente mediante el uso de toxinas en el proceso colonización del tejido vegetal (6,7,10). Por otro lado, las enfermedades causadas por patógenos biotróficos producen profundos cambios en la fisiología de los hospedantes, ya que alteran el metabolismo de las plantas al parasitar células vivas pudiendo producir reducciones en el área y capacidad fotosintética de las hojas, aumentos de la tasa respiratoria, la transpiración, consumo de asimilados y retención de nitrógeno por parte del patógeno, reducción en la turgencia celular y en la tasa de translocación de asimilados de los órganos afectados (2,14,16) pudiendo ser estos procesos los que implican las reducciones en la EUR evidenciadas en este trabajo.
La aplicación de fungicida provocó aumentos en los parámetros relacionados a la producción de biomasa y rendimiento (IAFV, DAFV, RFAa). En este sentido, su inclusión ha sido asociada con incrementos de rendimiento debido al mantenimiento de la fotosíntesis del cultivo durante el periodo de llenado del grano (12) y a que evitan el consumo de fotoasimilados por parte del patógeno. Algunos trabajos (6) hallaron que el uso de fungicidas para el control de enfermedades foliares en trigo está asociada a aumentos en el rendimiento en grano, peso específico del grano y PMG. Verreet & Klink (18) demostraron que gran parte de los carbohidratos de los granos provienen de la hoja bandera y sostienen que las hojas superiores del canopeo son las que más contribuyen al rendimiento del cultivo por lo que el aumento de la DAFV se correlaciona estrechamente con la acumulación de materia seca en el grano. El efecto que las enfermedades evaluadas ejercieron sobre sobre la tasa de crecimiento del cultivo alrededor de antesis, etapa crucial para la definición del rendimiento (3), produjo reducciones en el NGE y consecuentemente, en el NGT. Por otro lado, caídas en el PMG han sido vinculadas a infecciones de patógenos en post-floración que provocan reducciones en la DAFV o en la RFAa durante el llenado de los granos al afectar principalmente la tasa de llenado de granos (8, 12, 17).
CONCLUSIONES
Los resultados indican que Py.tritici-repentis (necrotrófico) afecta principalmente la absorción de radiación (mayores caídas en el IAFV y DAFV) a partir de mayores niveles de necrosis y senescencia foliar, en tanto que P.triticina (biotrófico) afecta más la EUR, probablemente por la interacción particular que este patógeno presenta con las células del hospedante. Los efectos detrimentales de los patógenos sobre estas variables afectan el rendimiento mediante disminuciones en el NGT, sustentadas por caídas en el NGE debido a reducciones en la tasa de crecimiento del cultivo en el periodo crítico, y por caídas en el PMG a partir de disminuciones en la tasa de llenado de granos.
BIBLIOGRAFÍA
1 Bancal, M.O; Robert, C; Ney, B. 2007. Modelling wheat growth and yield losses from late epidemics of foliar diseases using loss of green leaf area per layer and pre-anthesis reserves. Ann. Bot. 100, 777-789. [ Links ]
2 Boote, K.J; Jones, J.W; Mishoe, J.W; Berger, R.D. 1983. Coupling pests to crop growth simulators to predict yield reductions. Phytopathology 73, 1581-1587. [ Links ]
3 Borrás, L; Slafer, G.A; Otegui, M.E. 2004. Seed dry weight response to source-;sink manipulations in wheat, maize and soybean: a quantitative reappraisal. Field Crops Res. 86, 131-146. [ Links ]
4 Bryson, R.J; Paveley, N.D; Clark, W.S; Sylvester-Bradley, R; Scott, R.K. 1997. Use of in-field measurements of green leaf area and incident radiation to estimate the effects of yellow rust epidemics on the yield of winter wheat. Eur. J. Agron. 7, 53-62. [ Links ]
5 Carretero, R; Serrago, R.A; Bancal, M.O; Perelló, A.E; Miralles, D.J. 2010. Absorbed radiation and radiation use efficiency as affected by foliar diseases in relation to their vertical position into the canopy in wheat. Field Crops Research 116, 184-;195. [ Links ]
6 Dimmock, J.P.R.E; Gooding, M.J. 2002. The effects of fungicides on rate and duration of grain filing in winter wheat in relation to maintenance of flag leaf area. J. Agr. Sci., 138, 1-16. [ Links ]
7 Gooding, M.J; Dimmock, J.P; France, R.E; Jones, J. 2000. Green leaf area decline of wheat flag leaves: the influence of fungicides and relationships with mean grain weight and grain yield. Annals of Applied Biology 136, 77-;84 [ Links ]
8 Ishikawa, S; Hare, M.C; Kettlewell, P.S. 2011. Effects of strobilurin fungicide programmes and fertilizer nitrogen rates on winter wheat: severity of Septoria tritici, leaf senescence and yield. The Journal of Agricultural Science. pp. 1-16 [ Links ]
9 Miralles, D. J; Slafer, G.A. 1990. Estimación del área foliar en trigo: Generación y validación de un modelo. 11vo Congreso Nacional de Trigo. Pergamino. Cap.l, 76-85. [ Links ]
10 Ney, B; Bancal, M.O; Bancal, P; Bingham, I. J; Foulkes, J; Gouache, D; Paveley, N; Smith, J. 2013. Crop architecture and crop tolerance to fungal diseases and insect herbivory. Mechanisms to limit crop losses. Eur J Plant Pathol 135:561-;580 [ Links ]
11 Oerke, E.C; Dehne, H.W. 1997. Global crop production and the efficacy of crop protection -; current situation and future trends. Eur. J. Plant Pathol. 103,203-215. [ Links ]
12 Pepler, S; Gooding, M.J; Ellis, R.H. 2006. Modelling simultaneously water content and dry matter dynamics of wheat grains. Field Crops Res. 96, 46-93. [ Links ]
13 Robert, C; Bancal, M.O; Ney, B; Lannou, C. 2005. Wheat leaf photosynthesis loss due to leaf rust, with respect to lesion development and leaf nitrogen status. New Physiologist 165, 227- 241. [ Links ]
14 Schierenbeck, M; Fleitas, M.C; Simón, M.R. 2014. Componentes ecofisiológicos involucrados en la generación de biomasa afectados por enfermedades foliares en trigo. Revista Agronómica del Noroeste Argentino 34 (2), 247-250. ISSN: 0080-2069 y 2314-369X [ Links ]
15 Schierenbeck, M; Fleitas, M.C; Golik, S.I; Simón, M.R. 2015. Dinámica de área foliar verde y producción de biomasa aérea frente a infecciones de mancha amarilla y roya de la hoja. Revista Técnica de Cultivos invernales AAPRESID 2015. 22, 53-61. ISSN: 1850-0633 [ Links ]
16 Serrago, R.A; Carretero, R; Bancal, M.O; Miralles, D.J. 2009. Foliar diseases affect the ecophysiological attributes linked with yield and biomass in wheat (Triticum aestivum L.). European Journal of Agronomy 31,195-203. [ Links ]
17 Serrago, R.A; Carretero, R; Bancal, M.O; Miralles, D.J. 2011. Grain weight response to foliar diseases control in wheat (Triticum aestivumL.).Field Crops Research120, 352-;359. [ Links ]
18 Verreet, J.A; Klink, H. 2002. The biology of fungal pathogens. Vol. 1: Fungal pathogen and diseases of cereals. Department of Plant Pathology, Christian-Albrechts University. Kiel, Germany [ Links ]
19 Voegele, R.T; Mendgen, K.W. 2011. Nutrient uptake in rust fungi: How sweet is parasitic life?. Euphytica 179, 41-55 [ Links ]
20 Waggoner, P.E; Berger, R. 1987. Defoliation, Disease and Growth. Phytopathology 77, 393-398. [ Links ]
21 Zadoks, J.C; Chang, T.T; Konzak, C. F. 1974. A decimal code for the growth stages of cereals. Weed Research 14,415-;421. [ Links ]

