











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
El hongo fitopatógeno Verticillium dahliae Kleb, ocasiona la marchitez del olivo (Olea europea L.), enfermedad que causa graves daños al cultivo, pudiendo llegar a producir defoliación, muerte de ramas y plantas (14; 25; 3). Esta enfermedad se encuentra ampliamente distribuida en todas las regiones productoras de olivo a nivel mundial (26; 24) ocasionando considerables pérdidas económicas (19). Este hongo de suelo, causa graves daños en el cultivo de algodón (Gossypium hirsitum) (6) como así también en especies frutales, forestales, hortícolas y en malezas, produciendo marchitez localizada (14). Las malezas tienen un papel epidemiológico considerable, pueden actuar como reservorio de inóculo del hongo, manteniendo o aumentando el nivel en el suelo y contribuyendo a la dispersión del patógeno (14). V. dahliae crece en oscuridad, a pH neutros a alcalinos (pH 6-9) (14) se desarrolla y coloniza el xilema de plantas, produciendo micelio y conidios que son traslocados por la corriente transpiratoria, la que facilita el crecimiento fúngico aéreo (8). El hongo posee estructuras vegetativas de supervivencia (microesclerocios), que se producen en plantas senescente o tejidos muertos (24) pudiendo persistir en el suelo aproximadamente 15 años (8; 14; 15), permitiendo la dispersión y epidemiología (26). El largo periodo de supervivencia de los microesclerocios, el amplio rango de hospedantes que posee y el crecimiento parasítico en el xilema, obstaculiza la accesibilidad de productos químicos, lo que dificulta su control (13). Si bien la aplicación de productos químicos es la manera tradicional para el control de enfermedades en cultivos (11), ha sido cuestionada por los efectos que genera sobre el ambiente, la salud humana y la resistencia que desarrollan los microorganismos. Por lo que es necesaria la búsqueda de alternativas que sean eficientes y amigables con el ambiente (4; 11). El Control Biológico (C.B.) es sostenible, inocuo con el medio ambiente y compatible con otras herramientas para realizar un sistema integrado de manejo de enfermedades (14). El éxito de la estrategia depende de un equilibrio complejo de interacciones entre el patógeno, la planta huésped, el antagonista y el medio ambiente (14). Teniendo presente lo mencionado por Ezziyyani et al. (7), una de las condiciones del C.B. es que deben utilizar microorganismos antagónicos específicos contra el patógeno y que no ocasionen daño a la planta cultivada.
El hongo filamentoso Trichoderma es uno de los más utilizados como antagonista de fitopatógenos (29), ya que cuenta con múltiples mecanismos como la competencia por nutrientes y espacio, antibiosis y micoparasitismo, pudiendo también promover el crecimiento de las plantas y la activación de las respuestas de defensa de éstas (11; 23). Otra característica importante que presenta Trichoderma, es que posee fácil y rápido crecimiento y reproducción, en diferentes medios de cultivos (18) y sustratos, con rango de pH de 5.5 a 8.5 (18). Es viable su desarrollo en diversos medios artificiales: líquido, sólidos o semisólido, pudiéndose utilizar residuos agroindustriales, como por ejemplo granos de cereales y arroz; siendo este último el más utilizado para la producción de Trichoderma spp. a partir de procesos de fermentación artesanal e industrial (1).
Diversos estudios han demostrado que Trichoderma es un antagonista efectivo contra hongos fitopatógenos (21), ya que limitan el desarrollo de hongos dañinos como Rhizoctonia solani; Fusarium oxysporum y V. dahliae (11; 23).
El objetivo del presente trabajo es evaluar in vitro la capacidad antagónica de cepas nativas, de Trichoderma spp. frente al fitopatógeno V. dahliae, en diferentes condiciones de incubación del cultivo.
MATERIALES Y MÉTODOS
Muestreo y aislamiento del fitopatógeno: Se tomaron muestras de ramillas con síntomas característicos de Verticilosis, de una plantación de olivo, en el Dpto. Capayán, provincia de Catamarca (28°28’00 S; 65°47’00 O). Las muestras fueron recolectadas, siguiendo el protocolo sugerido por González Vera et al. (10), y luego fueron conservadas a 6°C. Posteriormente se realizó el aislamiento de V. dahliae siguiendo la metodología sugerida por Roca (22). Se cortaron las ramitas de entre 20 a 30 cm., retirando las hojas, posteriormente se lavó fuertemente con esponja y detergente diluido y enjuagó. Luego se desinfectó con hipoclorito de sodio 2 %, durante 2 min. Se enjuagó bajo el chorro del grifo por 45 - 60 min. Se continuó en cámara de flujo laminar con el descortezamiento superficial de las ramitas, realizado con bisturí. Se cortaron segmentos de material vegetal (cortex - xilema) de 5 cm., que fueron desinfectados con hipoclorito de sodio 1 %, por 15 s, enjuagados en agua destilada estéril y secados sobre papel tipo tissué estéril. Se continuó con la siembra de 5 a 6 trozos del material vegetal por placas de Petri con agar dextrosa papa (ADP) estéril e incubadas en oscuridad a 25 ± 1°C (26; 24) durante 14 días. Aislamiento de antagonistas: En los meses de septiembre, octubre y abril, se tomaron muestras de suelo y georreferenciaron 58 sitios diferentes (cultivados y no cultivados) de la provincia de Catamarca, siguiendo la metodología planteada por Vargas Gil et al. (28). Las muestras fueron acondicionadas y llevadas al laboratorio de fitopatología de la Facultad de Cs. Agrarias (UNCa.), donde fueron procesadas y luego utilizadas como inóculo en el sustrato. Preparación del sustrato y siembra de suelo: Los sustratos empleados fueron orujos de vid tinta y blanca, que se hidrataron durante 24 h y posteriormente se los escurrió y colocó en frascos de vidrio de 400 ml., completando el volumen de estos en un 75 %. Los frascos se taparon con papel de aluminio y se esterilizaron en autoclave con vapor saturado a presión a 1 atm., 121°C y durante 20 min. (1). Cada muestra de suelo fue sembrada por triplicado, en cámara de flujo laminar, espolvoreando 20 g de suelo sobre el sustrato estéril. Posteriormente se tapó con el papel de aluminio y agitó, llevando a incubación con fotoperíodo de 16 h de luz y 26 ± 1°C, durante 7 a 10 días. Se realizaron observaciones periódicas hasta la aparición de colonias típicas de Trichoderma, se corroboraron las características de estas con ayuda de microscopio estereoscópico Olympus SZ 11 - CTV y posteriormente se confirmó la presencia de conidios y micelio característicos del género mediante la observación con microscopio óptico Olympus BH-2 a 40 X de aumento (9). Los frascos que presentaron contaminación fueron desechados, quedando seleccionados aquellos que presentaron solo colonias características de Trichoderma. Aislamiento y conservación de cepas de Trichoderma spp.: Se tomó el inóculo contenido en el orujo y se realizaron aislamientos por agotamiento en superficie con siembras en estrías en superficie en medio de cultivo ADP y se incubó con fotoperíodo de 16 h de luz/ 8 h de oscuridad, a 26 ± 1°C. Las cepas aisladas fueron identificadas morfológicamente a nivel de género por medio de microscopio óptico, con aumento 40 X. Luego fueron conservadas en tubos de ensayo pico de flauta con ADP a 5°C. Identificación Molecular: Las cepas nativas con potencialidad antagonista y el fitopatógeno, se enviaron al laboratorio Macrogen - Corea donde se realizó la extracción del material genético, la amplificación de las regiones ITS1 - ITS4, con los primer: ITS1: 5´ (TCC CTA GGT GAA CCT GCT G) 3´ y ITS4 5´(TCC TCC GCT TAT TGA TAT GC) 3´ (11; 20; 29). Para la secuenciación se utilizó el kit: Big Dye terminator cycle sequencing Kit v 3.1, en el secuenciador Applied Biosystems model 3730XL, automated DNA sequencing system de Macrogen. Con estos resultados se procedió a la identificación molecular, mediante comparaciones de las secuencias obtenidas con la base de datos del GenBanK, espeficamente tomando como referencia la base de datos ARNr/ITS para hongos, la cual contiene registros curados y reanotados de las secuencias de la region ITS específicas del grupo Fungi. Las muestras secuenciadas, para cada cepa, fueron identificadas con la ayuda del software BlastN Report. Además, se calculó el porcentaje de similitud de las secuencias obtenidas con la cepa patrón, utilizando herramientas disponibles en la página del NCBI (National Center for Biotecbology Information). Preparación de cultivos madre: Los cultivos se realizaron en placas de Petri con cuatro cepas de Trichoderma spp. y el fitopatógeno V. dahliae. Se seleccionaron dos cepas de Trichoderma spp. que presentaron mejores características, para evaluar la capacidad antagónica contra el fitopatógeno V. dahliae. Se prepararon cultivos madres de las cepas en placas de Petri con ADP estéril, se incubaron a 25 ± 1°C con fotoperíodo de 16 h de luz para las cepas de Trichoderma spp. y en oscuridad para V. dahliae, durante cinco días para las primeras y de 12 días para V. dahliae. Ensayo de cultivo dual - (Bioensayo N° 1): Este ensayo se realizó en dos oportunidades, replicado en idénticas condiciones, con cinco repeticiones cada uno. Se efectuó un ensayo con un diseño completamente aleatorizado con arreglo factorial 3 x 2 x 2, un factor de tres cepas, el fitopatógenos y dos cepas antagonistas (M5A y TT), un segundo factor de dos pH distintos de 6.5 y 4,1 (acidificado con ác. láctico) y el tercer factor de 8 y 16 h. de luz. Los ensayos duales se prepararon en placas de Petri con ADP estéril, con dos pH de 6.5 y de 4.1 y se enfrentaron las cepas con potencialidad antagonista, contenidas en disco de ADP de 5 mm de diámetro con el fitopatógeno V. dahliae. Cada tratamiento conto con un testigo que consistió en un disco de ADP de 5 mm de diámetro del fitopatógeno. Posteriormente se incubó en condiciones controladas con fotoperiodos de 8 h de luz/16 h de oscuridad y otro de 16 h de luz/8 h de oscuridad (T1 y T2 respectivamente) a 26 ± 1°C durante cinco días. La acción antagónica se evaluó diariamente, mediante tres parámetros: 1) Tiempo de Contacto (T.C.) Antagonista - Patógeno, es el número de días en el cual ambas cepas entran en contacto. 2) Clase, es la competencia de sustrato a través de la Escala de Bell et al. (2), citado por Martínez et al. (18). 3) Porcentaje de inhibición del crecimiento (PIC), determinado con el crecimiento radial del fitopatógeno, PIC= [(C1 - C2) / C1] x 100; siendo C1: el crecimiento radial del fitopatógeno, testigo y C2: el crecimiento radial del fitopatógeno, en el cultivo dual. Lo que define el efecto inhibitorio, tomando el porcentaje de inhibición del crecimiento calculado a los 5 días. El crecimiento diario (C1 y C2) se midió sobre la recta que marca los tres cm. de distancia entre ambos microorganismos. Según la técnica propuesta por Reyes Ramírez et al. (21). Los datos obtenidos fueron analizados estadísticamente con el software estadístico Infostat (5). Microcultivos - (Bioensayo N° 2): Se determinó la acción micoparasitica de las cepas antagonistas seleccionadas, frente al fitopatógeno. Se efectuó un ensayo con un diseño completamente aleatorizado con arreglo factorial 2 x 2 x 2, un factor de dos cepas antagonistas (M5A y TT), un segundo factor de dos pH diferentes de 6.5 y de 4,1 y el tercer factor de 8 y 16 h de luz. Los microcultivos se acondicionaron en cuatro placas de Petri de 15 cm, en cámara húmeda. Posteriormente se incubo en condiciones controladas con temperatura de 25 ± 1°C y en las condiciones de iluminación establecidas. Se realizaron evaluaciones diarias para determinar el tipo de parasitismo que se presentaban, mediante observaciones con microscopio óptico a un aumento de 40 X. Según la técnica de Martins Corders y de Melo (17).
RESULTADOS Y DISCUSIÓN
Aislamiento del fitopatógeno: Se aisló V. dahliae de una planta de olivo, variedad Arauco (Cuadro N° 1). Aislamiento de antagonista: Se aislaron un total de 19 cepas de Trichoderma spp. Las que fueron observadas e identificadas con microscopio óptico (aumento 40 X). Mediante ensayos preliminares de crecimiento y actividad antagónica se seleccionaron las cepas M5A y TT para realizar los bioensayos, siendo la primera una cepa nativa y la última una cepa patrón, procedente del cepario de la Cátedra de Fitopatología de la Facultad de Ciencias Agrarias, Universidad Nacional de Catamarca (FCA - UNCa). Identificación Molecular: Se identificaron cinco cepas nativas con potencialidad antagónica, pertenecientes al género Trichoderma y a las especies T. hamatum, T. asperellum y T. longibranchiatum, de las que fueron seleccionadas solo dos por presentar mejores características; mientras que el fitopatógeno se identificó como Verticillum dahliae (Cuadro N° 1). Las cepas muestran un alto porcentaje de identificación respecto a las bases de datos proporcionadas por el NCBI. Además, se puede observar la variacion del porcentaje de similitud de las diferentes cepas con la cepa patrón. (Cuadro Nº 1).
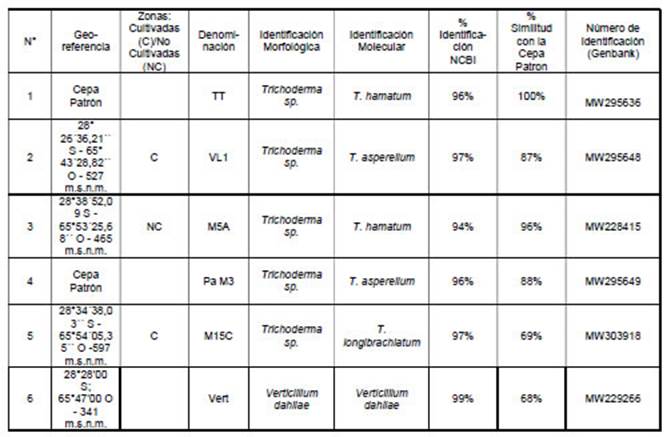
Tabla 1: Georeferencia de los aislamientos, Identificación Molecular y Número de Idenficación (Genbank) de cepas nativas antagonistas y del fitopatógeno.
Ensayo de cultivo dual (Bioensayo N°1): Los resultados muestran que, de acuerdo con el comportamiento presentado, en condiciones de pH 6,5 e incubación de 16 h de luz/8 h de oscuridad la cepa T. hamatum (TT) obtuvo el mejor TC al tercer día; mientras que con fotoperíodo de 8 h de luz/ 16 h de oscuridad el TC fue un 5,6 % mayor que el fotoperíodo de 16 h. luz/8 h. oscuridad. En tanto T. hamatum (M5A) tuvo un comportamiento de aproximadamente 15 y 10 % superior que TT con fotoperiodo de 16 h luz/8 h oscuridad, en los respectivos fotoperiodos. En cuanto a PIC, la mayor inhibición la presento TT con fotoperiodo de 8 h luz/16 h oscuridad, con un 35,33 %, siendo 10 % inferior con fotoperiodo de 16 h luz/8 h oscuridad; mientras que la cepa M5A, fue menos eficiente en un 40 % y 24 % en los respectivos fotoperiodos, respecto a la cepa TT 8 h luz/16 h oscuridad. Por lo que, en iguales condiciones de incubación, ambas cepas tienen comportamientos dispares. Respecto a la variable Clase, evaluada en los diferentes fotoperiodos, se clasificó como Clase 1 al quinto día logrando sobrepasar al fitopatógeno, lo que determina gran competencia por nutrientes y espacio por parte de las cepas antagonistas, logrando crecer y desarrollarse sobre la colonia del patógeno y cubrir el total de la superficie del medio de cultivo. El crecimiento y desarrollo de las cepas de Trichoderma no se vieron afectadas por las horas de luz, similar a los expresado por Valencia Abello y Castro Caicedo (27) quienes no obtuvieron diferencia entre los valores de pH de 4.5 y 6, en la concentración de inóculo de Trichoderma (espóras/ml. g de arroz) y las horas de luz tampoco afectaron el crecimiento. Según los resultados obtenidos, se evidenció mejor C.B. con la cepa TT, logrando mayor eficiencia que M5A, tanto en fotoperíodo de 16 y 8 horas de luz respectivamente, con pH de 6,5. Mientras que el pH de 4,1 se evidenció que no afecta el crecimiento de las cepas antagonistas evaluadas, en coincidencia con Valencia Abello y Castro Caicedo (27); en V. dahliae crecimiento fue nulo. Es importante destacar que en el crecimiento del fitopatógeno no se observó presencia de coloración negruzca, como indicio de presencia de microesclerocios. Por lo que a pH de 4,1 (con los dos fotoperiodos) la acción biocontroladora no pudo ser evaluada, caso contrario a lo planteado por Ezziyyani et al. (7) que en ensayo in vitro con T. harzianum frente a Phytophthora capsici, obtuvo mayor inhibición a pH ácidos. El pH es de vital importancia para el crecimiento de los vegetales, la disponibilidad de nutriente en el suelo y la actividad biológica. Por lo que regular el pH en el manejo del cultivo puede ser una estrategia para el manejo de las enfermedades causadas por V. dahliae, ya que el pH ácido (por debajo de 5.5) inhibe del crecimiento y la producción de microesclerocios y supervivencia. (14). De igual manera las condiciones de luz no afectan la actividad antagónica de Trichoderma, pero si regula la producción de conidios, siendo la oscuridad las condiciones óptimas para el crecimiento y desarrollo del patógeno (Cuadro N° 2).
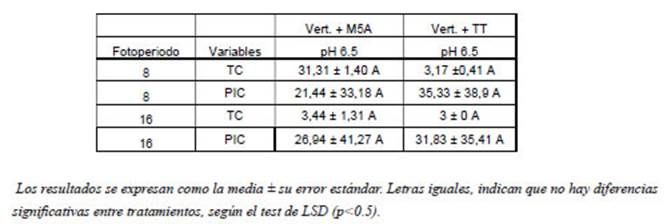
Tabla 2: Parámetros de antagonismo de V. dahliae, con fotoperiodos de 8 y 16 horas de luz y pH 6.5 del sustrato ADP.
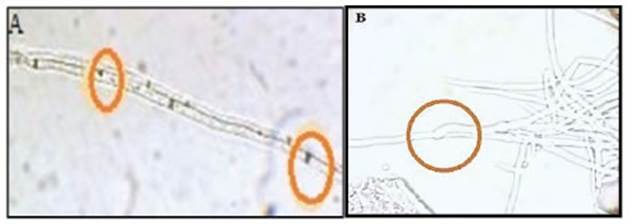
Microcultivos: El crecimiento y las interacciones antagónicas de las cepas TT (T. hamatum) y M5A (T. hamatum) variaron, según las condiciones de pH y fotoperiodos en el que se incubaron. La mejor interacción (crecimiento en paralelo, coilling, micoparasitismo y presencia de haustorios) la presentó la cepa M5A, con pH 4.1 y fotoperíodo de 8 h de luz/16 h de oscuridad. Mientras que, en ambos tratamientos de fotoperiodo, con pH 6,5 para las dos cepas antagónicas, se observaron 3 tipos de interacciones específicas (crecimiento en paralelo, coilling, micoparasitismo), por lo que se percibe que las condiciones de incubación no son limitantes para el proceso micoparasítico de estos antagonistas (Cuadro N° 3). En la Figura N° 1, se observa el parasitismo de la cepa TT en hifas de V. dahliae, fenómeno similar a lo encontrado en los otros tratamientos que se evaluaron. Por los resultados observados se presume que las cepas en estudio tienen capacidad antagónica en las diferentes condiciones de incubación evaluadas (fotoperiodo y pH), y presentan actividad lítica frente a V. dahliae logrando micoparasitarlo. En ambas cepas antagónicas se observó la colonización y producción de conidios sobre la colonia del fitopatógeno, por lo que se evidencia la capacidad de éstas para desarrollarse a expensas del contenido celular, sobre la cepa nativa de V. dahliae siendo eficientes controladores biológicos. Se estima que este proceso es coincidente con lo descripto por Infante et al. (12) y Maza et al. (16), quiénes señalaron que las cepas antagonistas crecen quimiotrópicamente en dirección al fitopatógeno, coincidente con M5A y TT frente a V. dahliae, se produce un reconocimiento entre Trichoderma y Verticillium, que al ser positivo las cepas de Trichoderma prosiguieron con la adhesión de hifas y coilling (o enrollamiento) a las hifas de V. dahliae. Martins Corder y de Melo (17) encontraron similares acciones antagónicas entre las cepas de Trichoderma sp. y V. dahliae, por lo que suponen que la reproducción de conidios de Trichoderma sobre V. dahliae se debe a que ciertos fitopatógenos estimulan la reproducción de los antagonistas. También se considera que la ausencia de microesclerocios de V. dahliae, puede deberse al desarrollo de conidios de TT sobre el fitopatógeno, con lo que el antagonista genera inhibición de la formación de estructuras de resistencia, como lo muestran Hernández Mendoza et al. (11). Sin embargo, a pH 4.1 y fotoperíodo de 16 h de luz/ 8 h de oscuridad, se observó la formación de verticilos y de conidios de V. dahliae.

CONCLUSIÓN
Las cepas TT y M5A presentan capacidad antagónica frente a V. dahliae, en condiciones de 8 h de luz / 16 h de oscuridad. Dicho fotoperíodo actúa como factor estimulante de la actividad antagónica, característica deseable y de interés para la agricultura sustentable, ya que el patógeno tiene crecimiento en condiciones de oscuridad. Las cepas M5A y TT seguirán siendo evaluadas con el objetivo de determinar su capacidad antagónica sobre plantines en invernáculo, con el objetivo de evaluar su incorporación a un manejo integrado de enfermedades del olivo.

