











 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
El efecto del cambio climático se observa actualmente en una amplia gama de ecosistemas y especies en todas las regiones del mundo en respuesta al aumento de la temperatura media (Rosenzweig et al., 2007).
El cambio climático ocasionó un aumento de la temperatura y se prevé que subirá más aun, lo cual causará pérdidas sustanciales en la productividad del cultivo de algodón debido al impacto negativo en los procesos fisiológicos de crecimiento y desarrollo (Al-Khatib y Paulsen, 1999).
Se espera que la temperatura global podría aumentar entre 1,4 y 5,8 ºC a finales del siglo xxi debido a los aumentos en las concentraciones de gases de efecto invernadero (IPCC, 2007).
El algodón en su estado nativo es un arbusto perenne que crece en un hábitat semidesértico y como tal requiere días cálidos (Gipson y Joham, 1969). Sin embargo, a pesar de que se origina a partir de clima cálido, sus rendimientos no logran valores óptimos bajo altas temperaturas (Oosterhuis y Snider, 2011).
En la mayor parte de las regiones productoras de algodón, las temperaturas actuales son ya cercanas o superiores a la temperatura óptima para su crecimiento y desarrollo. Por ello durante el periodo reproductivo es importante identificar y desarrollar técnicas de manejo que atenúen los efectos negativos de las altas temperaturas (Singh et al., 2004).
Temperaturas diurnas por encima de 30 °C se pueden presentar durante las etapas de floración y desarrollo del fruto en muchas regiones productoras de algodón en todo el mundo. Estas temperaturas elevadas pueden comprometer la fijación de órganos reproductivos, el rendimiento y los parámetros de calidad de la fibra (Reddy et al., 2004).
Diversos autores han definido las temperaturas que modulan el crecimiento y desarrollo del cultivo de algodón. Por ejemplo, la fase de primera flor a fin de floración efectiva presenta una temperatura base de 18 ºC, una temperatura óptima de 27 a 28 ºC y una temperatura crítica de 30 ºC (Reddy et al., 2004); mientras que la fase de maduración presenta una temperatura base de 20 ºC, una temperatura óptima de 25 a 30 °C y una temperatura crítica de 32 ºC (Bibi et al., 2003).
Zhao et al. (2005) observaron una mayor tasa de abscisión de las yemas florales y cápsulas jóvenes en las plantas cultivadas a temperaturas de 30 ºC durante el periodo reproductivo, en comparación con aquellas que crecen bajo temperaturas óptimas. Zeiher et al. (1995) demostraron que el número de cápsulas era bajo asociado con una temperatura mayor a 30 ºC, y que esta temperatura parece afectar específicamente el desarrollo de la capsula, ya sea por la destrucción del meristema reproductivo o por aborto del fruto (Hesketh y Low ,1968). En el mismo sentido, Morris (1964) demostró que la alta temperatura acorta el periodo de maduración, lo cual trae cápsulas con menos peso, acelera la precocidad y afecta la calidad de fibra del cultivo de algodón.
Jane et al. (2007) encontraron una interacción entre la temperatura y la arquitectura de la planta, así observaron que la temperatura de la canopia era superior en 5-6 ºC en surcos espaciados a 0,96 m en comparación con 0,34 o 0,17 metros. El sistema de surcos estrechos puede alterar la arquitectura de la planta y los procesos fisiológicos internos como consecuencia de la mayor densidad (Slosser et al., 1986).
El distanciamiento entre surcos influye en la arquitectura de la planta ya que estas pueden detectar la presencia de otras plantas debido a la sombra que generan (Ballaré y Casal, 2000). En el momento en que las plantas dentro de un cultivo empiezan a competir, el incremento de la densidad de plantas determina una reducción del tamaño de estas y un cambio en la disposición espacial (Smith, 1982; Casal et al., 1986; Ballaré et al., 1987). Las plantas sembradas en surco estrecho modifican el canopeo a través de la reducción de altura de la planta y mayor disponibilidad de luz en el medio y la parte superior de la canopia (Reta-Sanchez, 1992).
En este contexto, las prácticas de manejo agronómico, como el distanciamiento entre surcos, influyen en la arquitectura de las plantas, modificando el microclima y la temperatura del canopeo (Buxton et al., 1977) lo que nos permitiría lograr una adaptación significativa a los incrementos en temperatura que se vienen registrando en los últimos años en la región algodonera argentina. El objetivo de este estudio fue determinar la respuesta del cultivo de algodón al incremento de temperatura en el periodo reproductivo para dos espaciamientos entre surcos.
MATERIALES Y MÉTODOS
Sitio experimental
Los experimentos se realizaron a campo en la Estación Experimental Agropecuaria Sáenz Peña INTA (Instituto Nacional de Tecnología Agropecuaria), durante las campañas 2013/2014 y 2014/2015, ubicada en la localidad de Presidencia Roque Sáenz Peña, provincia del Chaco, Argentina.
El suelo donde se implantaron los ensayos corresponde a la serie Independencia y su clasificación taxonómica corresponde a un haplustol óxico que se encuentra en lomas tendidas que pertenecen a albardones fósiles sin evolucionar, de relieve normal. Suelo moderadamente profundo, la penetración efectiva de raíces de las plantas cultivadas puede llegar hasta alrededor de los 0,90 m, a partir de cuya profundidad hay un ambiente tóxico producido por altas concentraciones de sodio intercambiable (Ledesma et al., 1973).
Material vegetal
La variedad de algodón que se utilizó fue NuOpal RR la cual presenta excelente sanidad, es muy vigorosa, apta para suelos de baja a media fertilidad, ciclo largo (4600 grados días), tolerante a la enfermedad azul (cotton leafroll dwarf virus) y mancha angular (xanthomonas axonopodis pv. malvacearum). Dicha variedad se seleccionó en este trabajo por tener excelentes aptitudes agronómicas.
Aplicación de los tratamientos
Los tratamientos fueron una comparación entre dos distanciamientos entre surcos (1,00 m; D1 y 0,48 m; D2) con y sin generación de estrés térmico durante tres semanas consecutivas desde el inicio de floración. La duración de los tratamientos se basó en la duración de la etapa reproductiva más susceptible (Reddy et al., 1991).
La instalación de las carpas para generar el estrés térmico en la primera campaña (2013/14) se realizó el 23 de diciembre de 2013 (42 días después de la siembra; DDS; etapa de pimpollado) y en la segunda campaña (2014/15) se realizó el 29 de diciembre de 2014 (43 DDS).
Las carpas (una carpa por unidad experimental) que se utilizaron se fabricaron con una estructura de hierro con una altura de 2 m por 1,5 m ancho y 1,5 m de largo (superficie cubierta= 2,25 m2) de manera que puedan entrar los surcos en los cuales se realizó el registro. Se utilizaron cubiertas de un plástico transparente de 200 micras con los laterales abiertos a la altura de 1,20 m para permitir un óptimo intercambio gaseoso, evitar un excesivo calentamiento y permitir un crecimiento activo de planta (Cirilo y Andrade, 1996).
Al inicio de la floración se instalaron por repetición un data logger (Cavadevices) con 2 sensores que registraron la temperatura cada 15 minutos (un sensor se encontraba afuera de la carpa y el otro sensor en el interior de la carpa por debajo de planta y al costado del tallo) para poder registrar la temperatura durante todo el periodo (Neiff et al., 2016). Se calculó un índice de estrés térmico (sumatoria de tiempo térmico por encima de la temperatura umbral 30 ºC para el cultivo de algodón por hora / número de horas que la temperatura permaneció por encima de 30 ºC). En total se instalaron 16 sensores; 8 sensores adentro de las carpas (cuatro para distanciamiento 0,48 y cuatro para distanciamiento 1,00 m) y 8 sensores ubicados fuera de las carpas. Se consideró la duración del día (5:05-18:55 h) y duración de la noche (19:00-5:00 h).
Manejo de los experimentos
En la primera campaña (2013/14) la siembra se realizó el 11 de noviembre de 2013 y en la segunda (2014/15) fue el 17 de noviembre de 2014.
La siembra se realizó bajo siembra directa. A partir de los 10 días de producida la emergencia del cultivo se ralearon todas las unidades experimentales a 8 plantas por metro en las siembras distanciadas a 0,48 y a 16 plantas por metro en las siembras distanciadas a 1,00 m. De esta manera se obtiene la misma densidad en ambos sistemas de 160.000 plantas/hectárea.
Durante las dos campañas de ensayos (2013/2014) y (2014/2015) los tratamientos se regularon con Pix a una dosis de 300 cc/hectárea (para evitar el excesivo crecimiento vegetativo), se controlaron malezas y plagas según las recomendaciones de la sección algodón de la EEA-INTA Sáenz Peña. El ensayo se realizó en secano.
La cosecha se realizó el 21 de marzo para la primera campaña (2013/14) y el 27 de marzo en la segunda campaña (2014/15).
Mediciones realizadas a campo y en laboratorio
A los 30 DDS se seleccionó el área donde fue instalada la carpa y se realizaron los registros correspondientes.
En el inicio de floración (45 DDS) se instalaron las carpas durante 3 semanas seguidas. Luego de retiradas (50% de floración) las carpas, se registraron altura, número de nudos, número de pimpollos, de flores, de cápsulas, abscisión de órganos fructíferos (observando presencia o ausencia de frutos; cicatriz) y cápsulas abiertas de todas las plantas del metro lineal.
A los 80 DDS en todas las plantas de un metro lineal (8 plantas) de cada tratamiento se determinó el porcentaje de retención de órganos fructíferos (%ROF) mediante la fórmula [n.° órganos fructíferos retenidos / (n.º órganos fructíferos retenidos + n.º órganos fructíferos caídos)], dicha variable se cuantificó en porcentaje.
A los 95 DDS (posestrés) se midió el área foliar total por planta (AFP) con el Medidor Portable de Área Foliar LI-COR modelo LI-3000C. Las mediciones se hicieron sobre cuatro plantas por metro lineal en las cuatro repeticiones, totalizando 16 plantas por cada tratamiento. Las unidades de medida fueron en mm2 (área de scanner: 1 x 1 mm).
Para observar la disminución de radicación ocasionada por el plástico de la carpa, se midió la radiación incidente (µm/m-2/día-1) a las 12 am durante las tres semanas todos los días, tanto en el interior como fuera de la carpa sin el cultivo. Por única vez a los 95 DDS se midió la radiación incidente con un radiómetro (Cavadevices) con una barra de 1 metro de longitud. Se realizaron dos mediciones: 1.ª) en el estrato superior, colocando la barra por encima del ápice de la planta y 2.ª) en el estrato inferior, colocando la barra en el nudo número 6 sobre el tallo principal. La radiación interceptada (RDi) se calculó de la siguiente manera [1- (radiación inferior * radiación superior)]*100.
A los 102 DDS, y considerando que la apertura de cápsulas se encontraba por encima del 50%, se determinó precocidad (duración del ciclo) en cada tratamiento mediante el conteo del número de capullos (estructura fructífera abierta) y el número de cápsulas (estructura fructífera cerrada). La precocidad se calculó mediante la fórmula [N.º capullos / (N.º de capullos + N.º de cápsulas)]*100.
A los 102 DDS se procedió a la extracción de un metro lineal de las unidades experimentales (8 plantas) para la determinación de biomasa total y biomasa particionada, separando las estructuras vegetativas (tallo + hojas) y reproductivas (cápsula + pimpollo). Las muestras se llevaron a estufa a una temperatura de 60 ºC hasta peso constante. Se pesaron y se expresaron en g/m2.
A los 130 DDS se cosechó, se pesó el algodón en bruto (fibra + semilla) y luego se realizó el desmote (separación de la semilla de la fibra) mediante el empleo de una microdesmotadora eléctrica a sierras. El porcentaje de desmote es la relación entre [peso de fibra / peso fibra + peso semilla]*100. Las muestras de fibra se enviaron al laboratorio de calidad de fibra de la EEA Sáenz Peña para estimar los principales parámetros tecnológicos de la fibra: longitud (mm), resistencia (gr/tex) e índice de micronaire. Este análisis se realizó mediante un instrumento denominado HVI (High Volumen Instrument), marca: Uster y modelo: Spectrum.
Análisis estadístico
El diseño experimental que se utilizó fue de parcelas divididas (parcela principal: distanciamiento entre surcos y sub-parcela: estrés térmico) completamente aleatorizadas con 4 repeticiones (factorial: 2x2x4).
Las variables analizadas por ANOVA fueron: temperatura promedio horaria, área foliar, intercepción de la radiación, altura, retención de órganos fructíferos, precocidad, biomasa, componentes de rendimiento y parámetros de calidad de fibra mediante el empleo del paquete estadístico InfoStat/P versión 2017. Para la comparación entre medias se utilizó el test de Duncan (α=0,05). Se analizó la regresión lineal entre las variables de área foliar por planta y %ROF.
RESULTADOS
Descripción del ambiente
Precipitaciones
En la figura 1 se observan las precipitaciones acumuladas para ambas campañas. En el primer año de estudio se registró un total de precipitaciones de 561,1 mm, mientras que en el segundo un total de 443,1 mm. Ambos años presentaron un pico de máximas precipitaciones en noviembre donde se realizó la siembra manteniéndose las precipitaciones en torno a los 75 mm mensuales desde fines de diciembre (inicio de estrés térmico por altas temperaturas) a mediados de enero (finalización del estrés). Se destaca que en la segunda campaña el registro de precipitaciones fue más estable en los meses de enero, febrero y marzo.

Figura 1Precipitaciones mensuales durante el ciclo del cultivo para cada campaña (2013/2014) y (2014/2015) en Presidencia Roque Sáenz Peña (Chaco). Las flechas indican las fechas de siembra del cultivo de algodón y el inicio y finalización de estrés térmico por altas temperaturas en los meses correspondientes. Datos obtenidos de la Estación Meteorológica de la EEA Sáenz Peña.
Temperatura
Existieron diferencias estadísticamente significativas durante el día al analizar los valores de temperatura obtenidos tanto fuera como dentro de las carpas. Se observó este comportamiento en valores promedio y mediante un índice de estrés térmico. Con respecto a la temperatura nocturna no se observaron diferencias de temperatura entre los tratamientos, debido a que la carpa no se encontraba totalmente cerrada (tabla 1). Esto permitió una aireación constante y además la temperatura del aire a esas horas no era elevada.

Tabla 1Temperatura promedio en el interior y exterior de las carpas registrada mediante data loggers calibrados cada 15 minutos (5:05-18:55 h) y noche (19:00-5:00 h) del periodo de estrés evaluado para el cultivo de algodón. Índice de estrés térmico diurno dentro y fuera de la carpa. Durante las campañas 2013/14 y campañas 2014/15. Roque Sáenz Peña, Chaco.Letras distintas indican diferencias significativas (p<=0,05)
La aplicación de las carpas generó que a partir de las 10 de la mañana la temperatura en el interior comenzó a elevarse con respecto a la temperatura del ambiente, visualizando la diferencia entre interior y exterior (figura 2). La duración diaria de dicho estrés fue de 5 horas promedio durante tres semanas de la instalación de las carpas, haciendo un total de 105 horas promedio de estrés. La temperatura que se registró se mantuvo por encima de los umbrales óptimos (27-28 ºC) durante tres semanas continuas.

Figura 2Valores de temperatura media diurna (6:00,10:00,14:00, 16:00, 20:00) para dos distanciamientos entre surcos de algodón en el interior y exterior de las carpas (promedio de tres semanas y dos campañas) en Roque Sáenz Peña, Chaco.
En la figura 3 se observa que la temperatura de los tratamientos térmicos en ambos sistemas de siembra presentó diferencias respecto a la temperatura del día. El distanciamiento de 1,00 presentó diferencias estadísticamente significativas con respecto al distanciamiento de 0,48 m durante las horas del día 14:00 y 16:00 h, observándose un pico de máximas temperaturas alrededor de los 50 ºC (0,48 m) y 40 ºC (1,00 m). En distanciamientos de 0,48 m las diferencias de temperatura entre los tratamientos térmicos fueron menores comparadas con distanciamientos de 1 m.
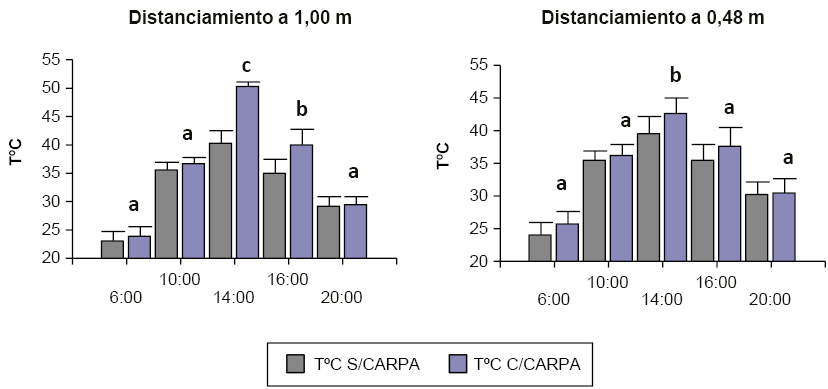
Figura 3Temperaturas en el interior y exterior de las carpas durante el periodo de estrés en ambas campañas (2013/14 y 2014/15) en algodón en Roque Sáenz Peña, Chaco para dos distanciamientos entre surcos (1,00 y 0,48 metros). El análisis estadístico individual para cada distanciamiento con su respetivo testigo.
Radiación incidente
La diferencia de incidencia de la radiación en el interior y exterior de las carpas sin la presencia del cultivo, es decir, la disminución de la radiación ocasionada por el plástico, fue de 10% de la radiación incidente.
Área foliar
Durante las dos campañas la variable área foliar total por planta (AFP) registrada en el periodo reproductivo (plena floración) presentó similar comportamiento. En los sistemas sometidos a estrés térmico el AFP fue superior a los testigos (sin estrés). En este sentido, la campaña 2013/14, la siembra a 1,00 m se diferenció estadísticamente de su testigo (sin estrés) en un 36,3%, a diferencia de la siembra a menor distanciamiento (0,48 m) que si bien incrementa un 17% su AFP con respeto a su testigo (sin estrés), no existen diferencias significativas. En la campaña 2014/15 se observaron resultados similares, los tratamientos con estrés térmico mostraron mayor AFP con una diferencia de 53 y 30,6% en siembras 1,00 y 0,48 m, respectivamente (tabla 2). Nuevamente, las diferencias significativas solo se observaron para los distanciamientos entre surcos de 1,00 m.
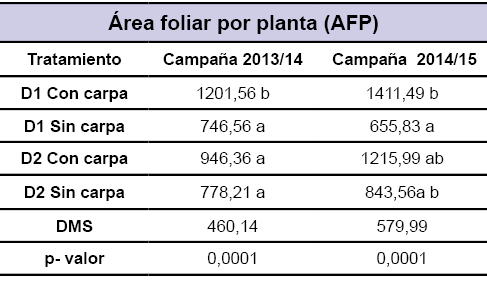
Tabla 2Área foliar por planta a los 95 DDS en los diferentes tratamientos durante las dos campañas.DMS representa la diferencia mínima significativa. Letras distintas indican diferencias significativas (p<=0,05).
Retención de órganos fructíferos
El porcentaje de retención de los órganos fructíferos a los 80 DDS mostró que los tratamientos testigos (sin estrés) presentaron mayor retención de órganos fructíferos con respecto a los tratamientos con estrés (tabla 3).
Cuando se compara la ROF entre ambos distanciamientos ante situaciones de estrés, se detectan diferencias estadísticamente significativas a favor de los surcos estrechos, observándose una menor disminución de los valores de la ROF para los D2 en ambas campañas (tabla 3). Cuando la comparación entre distanciamientos se hace sin estrés, existe claramente un efecto año ya que para la primera campaña no se presentan diferencias estadísticamente significativas, mientras que para la segunda campaña los D1 obtuvieron mayores valores de ROF que los D2 siendo esa diferencia estadísticamente significativa (interacción distanciamiento x año) (tabla 3).

Tabla 3Porcentaje de retención de órganos fructíferos (ROF) a los 80 DDS en los diferentes tratamientos (D1 y D2 con y sin estrés) en algodón en Roque Sáenz Peña, Chaco.DMS representa la diferencia mínima significativa. Letras distintas indican diferencias significativas (p<=0,05).
En la figura 4 se observa un análisis de regresión entre las variables AFP y ROF. En este se observa que existe un moderado grado de asociación entre ambas (R2 = 61).

Figura 4Análisis de regresión lineal del Área Foliar por planta (cm2) y porcentaje de retención de órganos fructíferos (%ROF) a los 80 DDS en función de los tratamientos (D1 y D2 con y sin estrés), (y= - 0,0743 +133,23, R2 =0,6176). Los puntos representan el promedio de ambas campañas en el cultivo de algodón en Roque Sáenz Peña, Chaco.
Precocidad
El comportamiento de la precocidad fue similar entre las dos campañas de estudio. Los tratamientos térmicos en ambos distanciamientos presentaron valores más altos de precocidad. En la campaña 2013/2014 los tratamientos térmicos presentaron diferencias estadísticamente significativas para ambos distanciamientos: en D1 la diferencia fue de 15,86% y para D2 fue del 11%; mientras que en la campaña 2014/2015 las diferencias fueron 15,7 y 8,6% en D2 y D1 respectivamente. Es decir, en ambos sistemas de siembra, la elevación de la temperatura acortó el tiempo de maduración (tabla 4). Comparando los tratamientos térmicos, D1 presentó mayor precocidad que D2 con diferencias estadísticas significativas entre dichos tratamientos (tabla 4).

Tabla 4Tiempo en que se produce la maduración de los frutos (precocidad) del cultivo de algodón expresada en porcentaje a 102 días después de la siembra en los diferentes combinación de tratamiento (D1 y D2 con y sin estrés) Roque Sáenz Peña, Chaco.DMS representa la diferencia mínima significativa. Letras distintas indican diferencias significativas (p<=0,05).
Biomasa seca aérea total
El análisis de los valores de biomasa total para las dos campañas mostró que ambos distanciamientos sometidos a estrés producen menos biomasa que sus respectivos testigos (tabla 5).
Para D1 las diferencias estadísticamente significativas fueron en ambas campañas y alcanzaron valores de biomasa total en el tratamiento térmico respecto a los testigos de un 22 y 26,4% menos para la primera y segunda campaña, respectivamente. En cuanto a D2, también presentó diferencias estadísticamente significativas para ambas campañas. En 2013/2014 la diferencia fue de 7,8% y en 2014/2015 de 17,3% menos de biomasa seca total para los tratamientos de estrés térmico con respecto al testigo (tabla 5). Comparando los tratamientos térmicos en la primera campaña las diferencias fueron significativas entre distanciamientos, donde D2 presentó 590 kg/ha más que D1; mientras que en la segunda campaña las diferencias no fueron estadísticamente significativas (tabla 5).

Tabla 5Biomasa seca total de algodón a los 127 días después de la siembra para todos los tratamientos analizados (D1 y D2 con y sin estrés) en las dos campañas en Roque Sáenz Peña, Chaco.DMS representa la diferencia mínima significativa. Letras distintas indican diferencias significativas (p<=0,05).
Rendimiento y sus componentes
Al analizar la variable rendimiento (kg/ha) se obtuvieron respuestas variables según las campañas, los tratamientos de distanciamiento y de estrés térmico.
La respuesta más importante se debió a las diferencias entre campañas, ya que en la campaña 2014/2015 el rendimiento fue prácticamente el doble del logrado en la campaña 2013/2014 para todos los distanciamientos y tipo de estrés, debido a que las precipitaciones fueron mayores y mejores distribuidas (tabla 6).
Cuando el análisis se realizó por distanciamiento, los resultados varían en función de la campaña considerada. Para la primera campaña, los tratamientos sin estrés rindieron más que los tratamientos con estrés: en D1 la reducción de los tratamientos con estrés fue de 25,8% y con el D2 de 18%. En la segunda campaña no se encontraron diferencias significativas dentro de cada distanciamiento entre los tratamientos térmicos, a pesar de presentar diferencias en valores absolutos favorables a los tratamientos sin estrés, siendo la reducción en D1 de 17% y en D2 de 11% (tabla 6).
Al igual que el rendimiento, el número de capullos fue muy superior en la campaña 2014/2015 que en la campaña 2013/2014 (tabla 6). En la campaña 2013/2014, el número de capullos por metro cuadrado solo presentó diferencias significativas con el D2, mostrando mayor número de capullos en los tratamientos sin estrés. En la campaña 2014/2015 se observaron diferencias significativas en los dos sistemas de siembras mostrando menor número de capullos en los tratamientos térmicos (tabla 6).
El peso de capullo mostró un comportamiento similar en ambas campañas, diferenciándose las respuestas principalmente por el distanciamiento. En los D1, los capullos fueron más pesados en el tratamiento sin estrés en relación con el estresado, originando diferencias significativas para la campaña 2014/15.

Tabla 6Rendimiento, peso individual de capullos y número de capullos de algodón en Roque Sáenz Peña, Chaco.DMS representa la diferencia mínima significativa. Letras distintas indican diferencias significativas (p<=0,05).
Rendimiento y parámetros de calidad de fibra
Durante las dos campañas no existieron diferencias estadísticamente significativas en el porcentaje de desmote y los parámetros de calidad de fibra (micronaire, longitud y resistencia) al comparar los diferentes tratamientos de distanciamientos con y sin estrés. (tabla 7).

Tabla 7Porcentaje de desmote (rendimiento de fibra) y principales parámetros de calidad de fibra de algodón en Roque Sáenz Peña, Chaco.egRegistros obtenidos del Laboratorio de Calidad de Fibra de la EEA de INTA de Presidencia Roque Sáenz Peña.Letras distintas indican diferencias significativas (p<= 0,05).
DISCUSIÓN
Las temperaturas altas son un fenómeno complejo que afecta a la fisiología, el crecimiento y la productividad en las plantas de algodón. Coincidiendo con Cicchino et al. (2010) y Neiff et al. (2016), el incremento de la temperatura durante el periodo reproductivo del cultivo de algodón mediante la utilización de las carpas de plástico trasparente fue efectivo, mostrando diferencias de temperatura durante el día. La elevación de la temperatura en el interior de las carpas provocó un estrés en el año donde las precipitaciones fueron menores, con cambios en el crecimiento y desarrollo, coincidiendo con Wahid et al. (2007) quienes reportaron que el estrés es causado por un ascenso en la temperatura máxima diaria por encima de un valor umbral y durante un periodo breve, lo que provoca daños irreversibles en el crecimiento y desarrollo de las plantas.
Al-Khatib y Paulsen (1999) mencionan que la magnitud de dicho efecto depende de características del estrés como intensidad, duración y tasa de incremento térmico. Precisamente, en este ensayo la temperatura que se registró se mantuvo por encima de los umbrales óptimos (30 ºC) durante tres semanas consecutivas, lo cual indicaría que la magnitud del efecto del estrés fue importante. En este marco, cuando las temperaturas fueron elevadas, el sistema de siembra distanciado a metro presentó mayor amplitud térmica, en comparación con el sistema de configuración en surcos estrechos, lo cual explica que a medida que se acorta la distancia entre surcos, las variaciones térmicas en el interior del cultivo son menores. Estos resultados coinciden con lo que expresan Jane et al. (2007) quienes encontraron una interacción entre la temperatura y la arquitectura de la planta, mostrando que la temperatura de la canopea era superior en 5-6 ºC en surcos espaciados a 0,96 m versus surcos espaciados a 0,34 o 0,17 metros. El efecto térmico causó mayor área foliar para ambos distanciamientos (mayor en el distanciamiento a metro) coincidiendo con Reddy et al. (1992) quienes encontraron que a temperaturas altas el área foliar era seis veces mayor que la de las plantas cultivadas a menor temperaturas, dado que la temperatura favorece el crecimiento. Constable (1975) encontró que los valores altos de área foliar afectan negativamente los rendimientos, causando abortos en las posiciones fructíferas, coincidiendo con los resultados de este ensayo, en los cuales los tratamientos sometidos a estrés presentaron mayor área foliar, pero menor rendimiento. El sistema de siembra distanciado a metro fue el que menor porcentaje de retención de órganos fructíferos presentó bajo condiciones de estrés térmico por altas temperaturas. En este ensayo, para ambos distanciamientos, hay una disminución del %ROF, pero esta merma no presentó diferencias estadísticamente significativas en siembras a menores distanciamientos (0,48 m), pero sí en las siembras de 1,00 m. Se destaca que el estrechamiento de surcos a 0,48 m amortiguó el efecto adverso del estrés térmico en el %ROF, no diferenciándose del testigo. Estos resultados concuerdan con lo que expresan Zhao et al. (2005) que observaron que cuando las temperaturas son elevadas en el periodo reproductivo hay mayor porcentaje de aborto.
Las plantas sometidas a estrés por alta temperatura mostraron valores más altos de precocidad respecto a las testigos en los dos distanciamientos entre surcos, lo cual significa que cumplieron su ciclo en menor tiempo generando, a su vez, una menor biomasa seca aérea total. Resultados similares fueron reportados por Morris (1964) que demostró que las altas temperaturas acortan el ciclo de cultivo. Yfoulis y Fasoulas (1978) expresan que las temperaturas altas acortan el tiempo de maduración del algodón por lo que sus cápsulas maduran antes y, como consecuencia, los valores de biomasa son significativamente menores, tal cual lo confirmaron Zeiher et al. (1994).
Con respecto al rendimiento (kg/ha), los tratamientos térmicos mostraron una reducción significativa en la primera campaña de estudio donde las precipitaciones fueron menores. Estos resultados coinciden con los reportados por Paytas et al. (2013) quienes expresan que las altas temperaturas afectan negativamente al rendimiento. Sin embargo, en la segunda campaña, donde las precipitaciones fueron más abundantes, al tener el cultivo un mayor suministro de agua el estrés térmico no se expresó por el enfriamiento evaporativo (Montaldi, 1995).
No se registraron diferencias significativas debido a los tratamientos térmicos en cuanto a los parámetros de calidad como ser los valores de longitud de fibra. En este último caso, las longitudes de fibra no llegaron a sus valores óptimos (29-30 mm) según lo expresado por Gipson y Joham (1969), donde expresan que la longitud máxima de fibra se obtiene cuando las temperaturas se encuentran entre 15 y 21 ºC.
Este conocimiento permitirá conocer el comportamiento de los sistemas de siembra ante los actuales y próximos cambios del clima, especialmente referidos a la elevación de la temperatura. Sin embargo, hace falta seguir investigando investigando sobre estrés térmico por altas temperaturas en diferentes fases del periodo reproductivo en distintos cultivares de algodón.
