











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Festuca alta, Schedonorus arundinaceus (Schreb.) Dumort [sinónimos: Festuca arundinacea Schreb., Lolium arundinaceum (Schreb.) S.J. Darbyshire, Schedonorus phoenix (Scop.) Holub], es una gramínea C3 perenne muy valorada en los sistemas de producción ganadera por su adaptación a un amplio rango de condiciones edafoclimáticas y su alto potencial de producción de forraje (Hannaway et al., 1999; Fribourg y Hannaway, 2007; Scheneiter et al., 2015). Al igual que otras gramíneas C3 puede establecer una relación generalmente simbiótica con hongos endófitos asexuales de transmisión vertical (Saikkonen et al., 1998). El endófito que infecta festuca alta, Epichloë coenophiala (Leuchtmann et al., 2014) ex Neotyphodium coenophialum Glenn, Bacon & Hanlin (= Acremonium Link Sección Albo-lanosa Morgan-Jones; Gams) (Glenn et al., 1996), desarrolla su ciclo de vida dentro de la planta hospedante, es asintomático y su dispersión tiene lugar únicamente a través de semillas infectadas, lo que se conoce como “transmisión vertical” (White et al., 1993; Clay y Schardl, 2002). Por un lado, la importancia agronómica de esta asociación está dada por los efectos positivos inducidos por la simbiosis pasto-hongo sobre el crecimiento y la tolerancia a estreses bióticos y abióticos (Malinowski y Belesky, 2000; White y Torres, 2009; Omacini et al., 2013) y por causar “festucosis”. Esta última hace referencia a distintas patologías que los alcaloides tóxicos producidos por el hongo provocan al ganado que consume plantas infectadas (Bacon et al., 1977; Hoveland, 1993; De Battista et al., 1995).
Por otro lado, se ha observado que pasturas de festuca alta sembradas con semilla libre de endófito son susceptibles a la invasión por plantas silvestres infectadas (ej.: resiembra natural); los mecanismos de contaminación son con semillas infectadas presentes en el banco o inmigración desde los bordes o campos aledaños, un factor clave en el avance de la infección endofítica (Shelby et al., 1989; Shelby y Schmidt, 1991; Shelby y Dalrymple, 1993; Barker et al., 2005). Estos mecanismos dependerán del tipo de pastura y del manejo de esta. Por ejemplo, se pueden mencionar el uso de cultivares de festuca libre de endófito o infectados con endófito amigable (Bouton et al., 2002; Gundel et al., 2013; Lugtenberg et al., 2016), el éxito reproductivo diferencial de plantas infectadas respecto a las libres de endófito (Gundel et al., 2008; 2011), la eficiencia de transmisión del endófito (Gundel et al., 2008; 2011), la aplicación de herbicidas, Defelice y Henning, 1990; Hill et al., 2010; Trevor et al., 2016), la intersiembra con leguminosas o gramíneas libre de endófito (De Battista et al., 1997; Tekeli y Ates, 2005; Petigrosso et al., 2018), el momento e intensidad de pastoreo (Evans et al., 2012) y el movimiento de los animales entre parcelas (Odriozola et al., 2002).
La transmisión vertical del endófito puede ser imperfecta, es decir, que las plantas infectadas pueden producir una proporción variable de semillas no infectadas, dependiendo de la especie o población en estudio y de las condiciones ambientales de crecimiento (Afkhami y Rudgers, 2008; Gundel et al., 2009, 2011; García Parisi et al., 2012). En este sentido, se sabe que la sequía y la aplicación de herbicidas disminuyen la transmisión vertical del endófito (Davitt et al., 2011; Gundel et al., 2011), pero poco se conoce sobre el efecto del pastoreo sobre esta. Por ejemplo, García Parisi et al. (2012) analizando el impacto del pastoreo (simulado mediante cortes y pisoteo) sobre la dinámica de transmisión del endófito E. occultans en Lolium multiflorum Lam. (raigrás anual) en microcosmos bajo diferentes contextos agroecológicos encontraron que la defoliación con pisoteo en dos momentos durante el ciclo de crecimiento (macollaje y encañazón) redujeron la producción de semilla y provocaron fallas en la transmisión del endófito respecto a los tratamientos no defoliados. Sin embargo, es escasa la información acerca de cómo operan los distintos factores ecológicos y de manejo agronómico en la regulación del reemplazo de plantas libres por plantas infectadas con endófito de tipo silvestre, que es común observar en las pasturas de la Cuenca del Salado (Colabelli et al., 2006; Petigrosso et al., 2013).
A partir de lo mencionado, y con la finalidad de desarrollar prácticas de manejo que permitan prevenir o disminuir la invasión de festuca infectada en pasturas libres de endófito, el presente trabajo tuvo como objetivo avanzar en el entendimiento de los factores que regulan dicha invasión. Para ello, se evaluó el efecto de dos frecuencias de defoliación sobre el crecimiento vegetativo y reproductivo de plantas de festuca alta de una población naturalizada e infectada con Epichloë coenophiala y de un cultivar comercial libre de endófito. Además, en las plantas infectadas se analizó el efecto de la frecuencia de defoliación sobre la transmisión vertical del endófito a las semillas.
MATERIALES Y MÉTODOS
Sitio y diseño experimental
El experimento se realizó en la Unidad Integrada Balcarce (Facultad de Ciencias Agrarias UNMdP–Estación Experimental Agropecuaria INTA Balcarce; 37° 45’ S, 58° 17’ O, 130 m s. n. m.) en micropasturas (unidades experimentales) establecidas sobre suelo tamizado y homogeneizado proveniente del horizonte A de un Argiudol típico (53 ppm de P Bray I, 4,3% MO y 30,1 ppm N‑NO3‑) en contenedores de 0,6 m de largo x 0,4 m de ancho x 0,2 m de profundidad ubicados al aire libre.
Se empleó semilla de festuca alta infectada con endófito silvestre (100% de infección, poder germinativo 82%) proveniente de un pastizal del partido de Mar Chiquita (37° 32’ S; 57° 55’ O) y semilla de festuca alta cultivar Continental TAITA, libre de infección (0% de infección, poder germinativo 87%), suministrada por GENTOS S.A. Los niveles de infección se corroboraron previo a la siembra empleando la técnica de Saha et al. (1988). Para ello, se realizó un pretratamiento que consiste en sumergir las semillas en hidróxido de sodio NaOH al 5% a temperatura ambiente por 12 h. Al cabo de este tiempo las semillas se enjuagaron con agua y se colocaron individualmente sobre un portaobjeto para separar las glumelas y proceder a su tinción (coloración directa) con una gota del colorante rosa de Bengala durante 2-3 minutos. Posteriormente se colocó un cubreobjeto y se observó al microscopio óptico Olympus CHK (400x) la presencia de hifas del hongo entre las células aleuroníferas del endosperma. Así, las semillas que contenían el endófito fueron diagnosticadas como positivas (E+) y las no infectadas como negativas (E-).
La siembra se realizó el 02/05/2016 ubicando 13 semillas a 5 mm de profundidad en cada uno de cuatro surcos distanciados 12 cm entre sí (52 plantas por contenedor, ej. 200 pl m‑2). Las fallas en la emergencia se subsanaron dentro de los 30 días después de la siembra, mediante el trasplante de plántulas extra, de tamaño similar, cultivadas paralelamente en almácigos. La fecha de emergencia promedio de las plántulas en las unidades experimentales fue el 25/05/2016, equivalente a un tiempo térmico desde la siembra de 144°Cd (temperatura base 4 °C; Colabelli et al., 1998; Agnusdei y Mazzanti, 2001).
El 20/10/16 se llevó a cabo un corte de limpieza para uniformar las micropasturas. A partir de esa fecha, se realizaron 8 cortes para los tratamientos con alta frecuencia y 4 cortes para los de baja frecuencia (tabla 1). La frecuencia de defoliación se estableció considerando el tiempo térmico acumulado (frecuencia alta cada 450±100°Cd y baja cada 850±100°Cd respectivamente). Los cortes se realizaron a 70 mm de altura desde el nivel del suelo con tijera de mano. El último corte se efectuó el 16/08/17 con el objetivo de permitir que las plantas pasen a estado reproductivo y evitar el corte de las varas florales.

Tabla 1Fechas de corte y tiempo térmico acumulado desde el corte de limpieza efectuado el 20 de octubre de 2016 (°Cd, temperatura base 4 °C) para dos frecuencias de defoliación: alta (AF, cada 450±100°Cd) y baja (BF, cada 850±100°Cd).
Se empleó un diseño experimental completamente aleatorizado con arreglo factorial. Los factores experimentales controlados fueron: tipo de pastura de festuca alta (FNE+: naturalizada e infectada con endófito silvestre, y FCE-: comercial libre de endófito) y nivel de frecuencia de defoliación (AF: alta, y BF: baja). En total se establecieron 20 contenedores experimentales: 2 niveles de defoliación x 2 niveles de infección endofítica x 5 repeticiones.
Determinaciones
Producción de biomasa aérea acumulada y número de macollos
Los cortes para alta y baja frecuencia de defoliación se realizaron en promedio a 470°Cd y 950°Cd, respectivamente, durante el período comprendido entre el 20/10/16 y el 16/08/17. La biomasa fresca se secó en estufa de aire forzado a 60 °C hasta peso constante y se pesó. En cada micropastura se identificaron 4 plantas de festuca alta en las que se contó el número total de macollos previo a la realización de cada corte.
Número de panojas, esfuerzo reproductivo, peso y número de semillas
En cada una de las 4 plantas marcadas se registró el número de macollos totales (reproductivos y vegetativos) para calcular el esfuerzo reproductivo (ej. panojas/macollos totales). Las panojas se cosecharon entre el 27/11/2017 y el 5/12/2017, cuando el raquis de estas presentaba un color amarillento. Se determinó el peso total de las semillas producidas en cada planta y, a partir de tres submuestras de 100 semillas se estimó el peso promedio de esas 100 semillas y así, con el dato de peso total, obtener el número de semillas por planta.
Determinación de la transmisión vertical del endófito
Se realizó el diagnóstico de la presencia del endófito en 20 semillas de cada una de las cuatro plantas identificada en cada micropastura empleando la técnica de Saha et al. (1988) previamente descripta. Con los resultados obtenidos se calculó el porcentaje de semillas infectadas en cada planta.
Datos meteorológicos y mantenimiento de los experimentos
El tiempo térmico acumulado se calculó utilizando los registros de temperatura del aire en casilla meteorológica a 1,50 m de altura de la Estación de Agrometereología de la EEA Balcarce del INTA, ubicada aproximadamente a 300 m del sitio experimental. Los contenedores se mantuvieron libres de malezas y con adecuada disponibilidad de agua durante el período experimental. Se aplicó nitrógeno (150 kg N ha‑1 año‑1 en forma dividida: 50 kg N ha‑1 a la siembra, 50 kg N ha‑1 luego del corte limpieza y 50 kg N ha‑1 después del último corte realizado previo a la encañazón). No se observaron ataques de plagas ni patógenos durante el desarrollo del experimento.
Análisis estadístico
Debido a la diferencia en el número de cortes entre ambos tratamientos de frecuencia de defoliación (ej. 8 y 4 valores para alta y baja frecuencia de defoliación, respectivamente), el cálculo de biomasa aérea acumulada con alta frecuencia de defoliación se efectuó sumando las cantidades cosechadas en cortes sucesivos de manera de obtener el valor de biomasa acumulada correspondiente al mismo período de acumulación de los tratamientos con baja frecuencia (ej. 4 valores en total para cada frecuencia de defoliación). Los efectos de los factores experimentales y su posible interacción se analizaron mediante ANOVA el cual, para las variables biomasa aérea acumulada y número de macollos, se efectuó con medidas repetidas en el tiempo. Se realizaron comparaciones de medias (LSD p<0,05) cuando se encontraron diferencias significativas (α=0,05). Debido a que de la variable respuesta número de macollo no cumplió con los supuestos de normalidad ni homogeneidad de varianzas debió transformarse (ej. ln del número de macollos). Todos los análisis se realizaron empleando el software estadístico R (R Development Core Team, 2016). Los modelos se ajustaron con la función lme del paquete nlme (Pinheiro et al., 2012). En la sección resultados se muestran como media ± error estándar (EE).
RESULTADOS
Producción de biomasa aérea acumulada y número de macollos
Al analizar la biomasa aérea acumulada de las micropasturas (figura 1) no se detectó interacción significativa entre el tipo de pastura, la frecuencia de defoliación y la fecha de corte (p=0,117), pero sí se encontró interacción significativa entre el tipo de pastura y la frecuencia de defoliación (p=0,009). En este sentido, se halló que con AF las plantas de FNE+ presentaron mayor acumulación de biomasa total que las de FCE- (168,19±8,84 y 149,12±3,74 g MS m-2 respectivamente), mientras que con baja frecuencia la situación fue inversa (FNE+: 104,13±7,07 y FCE-: 120,13±5,79 g MS m-2).

Figura 1Acumulación de biomasa aérea (g MS m-2, media ± EE, n=5) en micropasturas de festuca alta naturalizada en infectada con endófito silvestre (FNE+) y de un cultivar comercial libre de endófito (FCE-) bajo dos frecuencias de defoliación: alta (AF, cada 450±100°Cd) y baja (BF, cada 850±100°Cd). Los valores de AF corresponden a suma de dos cortes sucesivos (ver Materiales y métodos).
El número de macollos en las plantas de ambos tipos de pastura fue mayor con AF que con BF (p<0,0001) siendo en promedio 2346±302 y 1940±250 macollos m-2 respectivamente. El efecto tipo de pastura no fue significativo (p=0,061).
Número de panojas, esfuerzo reproductivo y número de semillas por planta
En la tabla 2 se muestran los resultados de las variables reproductivas. El número de panojas por planta fue mayor en FNE+ que en FCE- (2,85±0,20 y 1,55±0,34 panojas/planta, respectivamente, p=0,002) y no fue afectado por la frecuencia de defoliación (p=0,065). Se observó interacción significativa entre el tipo de pastura de festuca y la frecuencia de defoliación (p=0,0278) para el esfuerzo reproductivo. Así, si bien las plantas FNE+ tendieron a mostrar mayor esfuerzo reproductivo que las FCE-, la diferencia solo fue significativa en BF. Por último, la producción de semillas fue mayor en las plantas FNE+ que en las FCE- (316,44±35,03 y 183,53±18,19 semillas/planta, respectivamente, p=0,0033) y, aunque no fue afectada por la frecuencia de defoliación (p=0,2581), el aumento de esta tendió a disminuir el número de semillas producidas únicamente en las plantas FNE+.
Transmisión vertical del endófito
Todas las semillas analizadas producidas por las plantas FNE+ resultaron positivas al diagnóstico de endófito, por lo que no hubo efecto de la frecuencia de defoliación sobre la transmisión de este.
DISCUSIÓN
El presente experimento compara la respuesta a la frecuencia de defoliación de plantas de festuca alta provenientes de semilla mejorada libre de endófito y de una población naturalizada infectada con endófito silvestre con la finalidad de avanzar en el conocimiento de los mecanismos que explican el avance del porcentaje de infección en pasturas sembradas con semilla libre de endófito en la Cuenca del Salado (Petigrosso et al., 2013). Bajo estas estas condiciones experimentales no es posible evaluar el efecto aislado del endófito debido a las diferencias genotípicas existentes entre cultivares, poblaciones y aun dentro de la misma población (Hill et al., 1990; Saikkonen et al., 2004).
En nuestras condiciones experimentales se observó que la tendencia del número de macollos registrada (figura 2) coincide con la publicada por Parsons et al. (1983) y Chapman y Lemaire (1996) quienes hallaron que pasturas manejadas con alta frecuencia de defoliación desarrollaron una estructura con una mayor densidad de macollos de menor tamaño que aquellas sometidas a bajas frecuencias de defoliación, lo que se explicaría por la ley de compensación tamaño/densidad (Matthew et al., 1995, 2000). Similarmente, Petigrosso et al. (2018) encontraron que la densidad de macollos de festuca alta infectada creciendo en micropasturas puras y consociadas con Lotus tenuis tendió a ser mayor en alta frecuencia de defoliación que en baja frecuencia (alta: cada 7-9 días, y baja: cada 14-21 días).
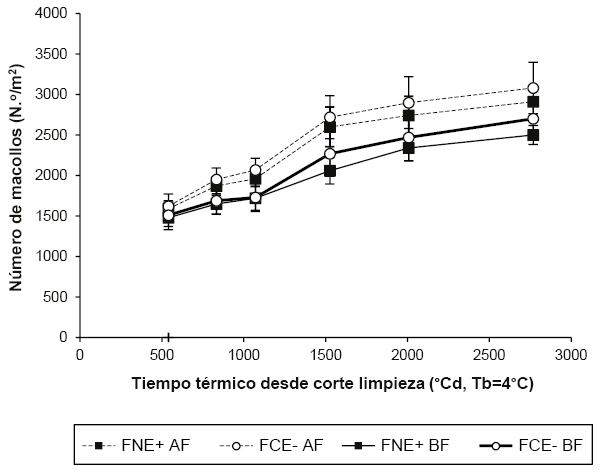
Figura 2Dinámica del número de macollos por planta (media ± EE, n=5) en micropasturas de festuca alta naturalizada en infectada con endófito silvestre (FNE+) y de un cultivar comercial libre de endófito (FCE-) bajo dos frecuencias de defoliación: alta (AF, cada 450±100°Cd) y baja (BF, cada 850±100°Cd).
Respecto a la transmisión vertical del endófito, a diferencia de los resultados obtenidos por García Parisi et al. (2012), no se registraron fallas de trasmisión del hongo de la planta madre a las semillas en ninguna de las frecuencias de defoliación aplicadas en este experimento. Ello podría atribuirse a que los autores antes mencionados trabajaron con una especie anual (Lolium multiflorum) infectada con endófito, con pastoreo simulado y sometida a solo dos cortes durante el ciclo de crecimiento en el tratamiento con defoliación. En ese sentido, se ha comprobado que tanto los disturbios directos e indirectos generados por la presencia de herbívoros sobre el suelo como las condiciones ambientales afectan la transmisión del endófito a la semilla (Bardgett et al., 2005; Davitt et al., 2011). Además, resulta interesante destacar que con BF las plantas FNE+ presentaron mayor esfuerzo reproductivo que las FCE- y tendieron a producir menor número de panojas y semillas/planta en AF que en BF (tabla 2). Estos resultados indican que las prácticas de manejo frecuentemente observadas en los festucales infectados de la Cuenca del Salado, que consisten en cerrar los lotes en primavera avanzada no son adecuados ya que favorecen la producción de semilla y la acumulación de esta en el banco del suelo, lo que conlleva a un aumento progresivo de la proporción de plantas infectadas (Castaño J., comunicación personal). Es por ello que una de las estrategias de manejo sugeridas para reducir la severidad de la festucosis consiste en someter a una alta frecuencia de defoliación a las pasturas de festuca alta E+ a fin de mantenerlas en estado vegetativo, que los animales consuman mayoritariamente láminas foliares, las cuales presentan menor contenido de ergoalcaloides (White et al., 1993) y, de este modo, disminuir la producción de semilla que es la principal fuente de contaminación y expansión de esta problemática.

Tabla 2.Número de panojas, esfuerzo reproductivo y número de semillas por planta (media ± EE) en festuca alta naturalizada en infectada con endófito silvestre (FNE+) y de un cultivar comercial libre de endófito (FCE-) bajo dos frecuencias de defoliación: alta (AF, cada 450±100°Cd) y baja (BF, cada 850±100°Cd).
Entre los aspectos que se deberían analizar en futuras investigaciones se pueden mencionar: la dinámica de avance de la infección endofítica en pasturas de festuca alta y pastizales en experimentos a campo y de larga duración, la competencia entre plantas infectadas y libres de endófito (con el mismo fondo genético), la dinámica del banco de semillas del suelo, el impacto del consumo preferencial de semillas y plántulas por herbívoros, el establecimiento de plantas infectadas y libres de endófito propagadas a través del excremento del ganado y la posible interacción con otros simbiontes a nivel de la rizósfera.
CONCLUSIONES
En función de los resultados obtenidos podemos concluir que, bajo nuestras condiciones experimentales, la frecuencia de defoliación alta favoreció la producción de biomasa aérea de las plantas de festuca de la población naturalizada e infectada con respecto al cultivar libre de endófito, sin embargo ocurrió lo contrario en las plantas sometidas a la frecuencia de defoliación baja. Independientemente de la frecuencia de defoliación, las plantas de la población naturalizada e infectada tendieron a producir mayor número de panojas y semillas que las del cultivar libre de endófito, aunque la producción de estas últimas tendió a disminuir con el aumento de la frecuencia de defoliación. No se registraron fallas en la transmisión vertical del endófito a la semilla producida por las primeras. Con base en estos resultados, sería recomendable aplicar una frecuencia de defoliación que, sin afectar la supervivencia de las plantas, sea lo suficientemente alta para permitir mantener la pastura en estado vegetativo y disminuir la producción de semilla (principal fuente de contaminación y expansión de festuca E+).
