










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
versão On-line ISSN 1851-2372
Bol. Soc. Argent. Bot. vol.54 no.1 Córdoba mar. 2019
Paleobotánica y Palinología - Paleobotany and Palynology
Análisis palinológico de superficie de hojas y tallos de ítems dietarios de Lama guanicoe (Fam. Camelidae) en Patagonia (Argentina): implicancias en la identificación del origen del polen en coprolitos
Palynological analysis of stems and leaves surface of food items of Lama guanicoe (Fam. Camelidae) in Patagonia (Argentina): implicances in the identification of the pollen source in coprolites
Nadia Jimena Velázquez1y Lidia Susana Burry2
1. CONICET-IPROSAM, Departamento de Biología, Facultad de Ciencias Exactas y Naturales, Universidad Nacional de Mar del Plata, Funes 3350, Mar del Plata, Argentina.
2. UNMdP. Departamento de Biología, Facultad de Ciencias Exactas y Naturales, Universidad Nacional de Mar del Plata, Funes 3350, Mar del Plata, Argentina.
*nvelazquez@mdp.edu.ar
Summary
Background and aims: One of the problems of copropalynology within palaeodiet analysis is the elucidation of the source of the pollen content which can be the result of pollen contamination of food or water or post-depositional pollen contamination. The aim of this study was to analyze the pollen spectrum on the surface of stems and leaves of Empetrum rubrum, Senecio flaginoides, Nardophyllum obtusifolium and Mulinum spinosum, food items of Lama guanicoe from Perito Moreno National Park, to evaluate the pollen contamination of food items.
M&M: Stems and leaves of E. rubrum, S. flaginoides, N. obtusifolium and M. spinosum were collected in Roble river valley and observed under scanning electron microscope. Then, acetolysis and pollen count under optic microscope were performed.
Results: The results showed that dominant pollen types, except in the sample of N. obtusifolium, come from the mother plant. However, it is significant the presence of other pollen types such as Nothofagus, Nassauvia, Podocarpus, Poaceae and Rumex on plants surface
Conclusions: The results of this study are related to the epidermic characteristics and provide relevant information about the origin of pollen within coprolites of Lama guanicoe to reconstruction of paleodiet.
Key words Pollen contamination, coprolites, Lama guanicoe.
Resumen
Introducción y objetivos: Una de las problemáticas de la copropalinología para estudios de paleodieta es identificar la procedencia del polen que podría deberse a la contaminación polínica del alimento, al beber agua con polen en suspensión, o a contaminación postdepositacional. El objetivo de este trabajo es analizar el espectro polínico de la superficie de tallos y hojas de Empetrum rubrum, Senecio flaginoides, Nardophyllum obtusifolium y Mulinum spinosum, que conforman la dieta de Lama guanicoe del Parque Nacional Perito Moreno, para evaluar la contaminación polínica del alimento.
M&M: Se recolectaron tallos y hojas de cada una de estas especies en el valle del río Roble. Hojas y tallos de cada ejemplar se observaron al microscopio electrónico de barrido y se realizó acetólisis y el recuento polínico bajo microscopio óptico.
resultados: Los resultados evidencian que los tipos polínicos dominantes, salvo en las muestras de N. obtusifolium, proceden de la misma especie. Sin embargo, es importante la presencia de otros tipos polínicos como Nothofagus, Nassauvia, Podocarpus, Poaceae y Rumex sobre la superficie de las plantas.
conclusiones: Los resultados de este estudio se relacionan con las características epidérmicas de las plantas y aportan información referente a la procedencia del polen en los coprolitos de Lama guanicoe, permitiendo precisar la información para reconstruir paleodietas.
Palabras claves Contaminación polínica, coprolitos, Lama guanicoe.
Introducción
El estudio polínico de heces actuales y coprolitos se ha orientado a la reconstrucción de la dieta de los organismos y particularmente la de los coprolitos también a la reconstrucción paleoambiental (Reinhard & Bryant, 1992; Carrión et al., 2001, 2004; Horrocks et al., 2003, 2008; Velázquez et al., 2010; Velázquez & Burry, 2012; Wood et al., 2012). Varios autores señalan que un punto importante en el estudio de coprolitos es dilucidar la procedencia polínica (Alcover et al., 1999; Carrión et al., 2004; Fernández-Jalvo et al., 2010). Por ejemplo, los granos de polen encontrados en los coprolitos de herbívoros y omnívoros tienen relación con su dieta y con la disponibilidad de las plantas en el ambiente, mientras que en carnívoros y también en omnívoros, se relacionan fundamentalmente a los granos que pudieran estar contenidos en el tracto digestivo de las presas consumidas (Carrión et al., 2001). Es así que la presencia del polen en los coprolitos de herbívoros y omnívoros puede ser el resultado de:
1) la dieta por la ingesta de hojas, tallos, fores y frutos con granos de polen que quedan adheridos en la superficie de estos órganos (contaminación polínica del alimento) (Bryant & Holloway, 1983).
2) la toma de agua donde se encuentran suspendidos granos de polen provenientes de la lluvia polínica o de plantas acuáticas.
3) la inhalación de aire.
4) la depositación de polen sobre la superficie de las heces luego de la deposición, lo que refeja la contaminación de las heces por polen con dispersión anemófla (Carrión et al., 2001, 2005). Esto ocurre porque una vez que el organismo depositó las heces, el mucus que las rodea, originado en la mucosa intestinal, actúa como una trampa para los granos de polen provenientes de la lluvia polínica (Chaves, 2000). Además, la concentración y preservación del polen en el interior de los coprolitos están infuenciadas por varios factores: a) el tipo de polinización de las plantas (zoófla, anemófla, hidrófila o autopolinización), b) la fisiología del sistema digestivo, c) el hábito alimenticio o comportamiento del organismo productor, d) variables ambientales como la temperatura, humedad, e) el grado de preservación del coprolito, etc.
Por esta razón para los estudios de dieta a partir del análisis de coprolitos es importante reconocer la procedencia de los granos, si fueron parte de la dieta o ingresaron por otra vía.
La emisión y depositación de polen consiste en la apertura de las fores o extensión de las anteras, la dehiscencia de las anteras, la depositación por gravedad sobre las partes forales o las hojas y tallos que se encuentran debajo de las fores, y la remoción del polen de los diferentes órganos por acción del viento o precipitaciones (D'Antoni, 2008).
Existe poca información sobre la adherencia de polen sobre diferentes plantas o partes de plantas y el efecto sobre el contenido de polen en los coprolitos. Tauber (1967) analizó el polen atrapado en la superficie de las ramas pegajosas y pubescentes del sauce y de las ramas de superficie lisa del abedul. Este autor recuperó más de 100.000 granos por gramo de todas las especies de la región en las ramas del sauce y solo algunos cientos de granos en las ramas de abedul. Con estos resultados se refeja el papel que juega la adhesividad de la superficie de las plantas que infuye en las interpretaciones sobre dieta a partir del análisis del contenido de polen en los coprolitos.
Moe (1983) analizó los espectros polínicos de heces de ovejas y de superficies de hojas de plantas consumidas por estos herbívoros, e infrió que el polen anemóflo de árboles cercanos al área de estudio se debería haber depositado sobre las plantas consumidas por las ovejas antes de que sean lavadas por las precipitaciones.
Por otra parte, O'Rourke (1991) analizó el espectro polínico de la superficie de hojas de diversas especies que crecen en el desierto de Sonora. Los resultados mostraron, por un lado, que las especies presentaban en mayor proporción polen de la planta madre y por el otro, que existían diferencias entre las diferentes especies en la abundancia de polen capturado que estarían relacionadas a las características epidérmicas de las plantas.
El guanaco, Lama guanicoe, (Müller, 1776) es un pseudo-rumiante, cuyo hábito pasteador y ramoneador explicaría en parte, su amplia distribución geográfica y la gran adaptabilidad para vivir en diversos ambientes (Wheeler, 1991). Presenta adaptaciones fisiológicas a condiciones áridas y tiene la capacidad de soportar el estrés hídrico, siempre y cuando el contenido de humedad de los vegetales consumidos sea lo suficientemente alto. Su hábito de alimentación es generalista, de selección intermedia (Raedeke, 1980; Franklin, 1982, 1983; Wheeler, 1991; Puig et al. 1996; Miñoz, 2008). Estos hábitos y sus características fisiológicas le posibilitan habitar pampas altas, muy secas y con muy baja calidad de forraje (Lauenroth, 1998). Su área de distribución comprende ambientes con marcadas diferencias en la estructura de la vegetación, como la estepa patagónica, los bosques subantárticos (Franklin, 1983) y el ecotono bosque-estepa. Se alimenta principalmente de los estratos herbáceo (Poaceae, Juncaceae y Cyperaceae) y arbustivo (Raedeke, 1978) y en algunos casos, del estrato arbóreo cuando migra hacia el interior del bosque durante el invierno (Bank et al., 2002). Posee un rango de acción amplio que varía de acuerdo a las condiciones del lugar y a la época del año, oscilando entre 1,30 y 9 km2 (Burgi, 2007).
Velázquez & Burry (2012) analizaron el espectro polínico de heces actuales de guanaco provenientes de diferentes estaciones del año en el valle del río Roble (Parque Nacional Perito Moreno, Santa Cruz). Los resultados mostraron diferencias en el espectro polínico que se relacionan a la estación de polinización y/o a variaciones en la dieta. Las autoras señalan que para evaluar si la concentración de tipos polínicos con dispersión anemófla en las heces corresponde a la dieta o son producto de contaminación polínica de las plantas consumidas, se debería analizar el contenido polínico de la superficie de tallos y hojas de plantas que representan items alimentarios del área de forrajeo de los guanacos.
El objetivo de este trabajo es analizar el espectro polínico de la superficie de tallos y hojas de Empetrum rubrum Vahl ex Willd., Senecio flaginoides DC., Nardophyllum obtusifolium Hook & Arn. y Mulinum spinosum (Cav.) Pers., especies que conforman parte de la dieta de Lama guanicoe en el Parque Nacional Perito Moreno (PNPM), Santa Cruz.
Los resultados de estos análisis serán útiles para la interpretación de los espectros polínicos de los coprolitos.
Área de Estudio
En la zona del PNPM se trabajó en el Cerro Casa de Piedra (CCP) (47º57´S; 72º05´O, 900 msnm), ubicado en una faja transicional entre el bosque cordillerano y la estepa patagónica, en la cuenca del río Roble y del Lago Burmeister. Entre los años 1930 y 1940 el bosque de Nothofagus sp. que se desarrolla frente al Cerro ha sido modificado por talas e incendios (Pardiñas, 1998).
La vegetación que se encuentra actualmente en los alrededores del CCP desde la base del cerro hacia el río Roble está dominada por una franja de Empetrum rubrum que forma un denso murtillar con Gaultheria mucronata; luego en algunos sectores aparece una estepa graminosa de Festuca pallescens; continúa un murtillar con Azorella monanthos y árboles adultos y renovales dispersos de Nothofagus pumilio; luego una estepa arbustiva con predominio de Nardophyllum obtusifolium, Mulinum spinosum, Senecio flaginoides y Adesmia boronioides. En la ribera del río Roble, se observa una estepa graminosa-arbustiva dominada por N. obtusifolium acompañada por gramíneas y Acaena sp. (Observación personal).
Materiales y metodos
En el valle del río Roble se recolectaron tallos y hojas de ejemplares de Empetrum rubrum "murtilla" (Ericaceae), Senecio filaginoides "mata mora" (Asteraceae subfam. Asteroideae), Nardophyllum obtusifolium "mata torcida" (Asteraceae subfam. Asteroideae) y Mulinum spinosum "neneo" (Apiaceae) (n=16), en una campaña realizada en abril de 2014.
Para la identificación de las especies se utilizó la Bibliografia de Correa (1971-1999) y Dimitri (1987). Además se consultó el Catálogo de Plantas Vasculares del Instituto Botánica Darwinion (http:// www.darwin.edu.ar/Proyectos/FloraArgentina/ fa.htm).
En el laboratorio se seleccionaron hojas y tallos de cada una de las especies y se montaron sobre tacos, luego fueron cubiertas con oro-paladio para la observación al microscopio electrónico de barrido (MEB). Se observaron las estructuras epidérmicas de los tallos y hojas de las cuatro especies para evaluar la adhesividad de la superficie del obstáculo en la eficiencia de retención de granos de polen.
Para la extracción polínica, se pesaron dos gramos, entre hojas y tallos, de cada ejemplar; se adicionó agua destilada y una tableta de esporas de Lycopodium clavatum (Batch N°124961, media= 12542 esporas/tableta) a cada una de las muestras y se fltraron a través de una malla de 260 μm.
Se realizó la extracción polínica, según D'Antoni (1979) y Faegri & Iversen (1989) y consistió en:
a) concentración del fltrado por centrifugación 2500 rpm por 5 minutos;
b) deshidratación con ácido acético puro.
c) eliminación del material celulósico mediante acetólisis (la mezcla acetolítica está formada por 9 partes de anhídrido acético y 1 de ácido sulfúrico) y posterior lavado con ácido acético puro para evitar la precipitación del octacetato de celulosa (D'Antoni, 1979). La mezcla acetolítica mediante hidrólisis remueve polisacáridos, como la celulosa, que están presentes en el citoplasma y en la superficie de los granos de polen (Erdtman, 1943; Faegri & Iversen, 1989). Por último, se concentró el residuo obtenido mediante centrifugación a 2500 rpm por 5 min y las muestras se almacenaron en tubos Kahn con el agregado de 2 gotas de glicerina.
Se realizaron preparados semipermanentes (D'Antoni, 1979). La observación microscópica se realizó utilizando un microscopio óptico binocular Olympus BH-2 con aumentos de 400X y 1000X. La identificación y el recuento polínico se llevaron a cabo utilizando Bibliografia especializada (Heusser, 1971; Markgraf & D'Antoni, 1978; Moore et al., 1991). Además, se utilizó la palinoteca de referencia del Laboratorio de Palinología y del Grupo de Palinología y Bioantropología (Universidad Nacional de Mar del Plata, Argentina). La nomenclatura de los tipos polínicos se estableció de acuerdo a Joosten & De Klerk (2002) y De Klerk & Joosten (2007). La suma polínica incluyó a todos los tipos polínicos y esporas, hasta alcanzar una suma de al menos 200 entre granos y esporas.
El recuento polínico se expresó en porcentaje y concentración (n° de granos/g de muestra). Los resultados de porcentajes se presentaron en un diagrama polínico porcentual (Fig. 5) y los de concentraciones en una tabla (Tabla 1).
resultados
Observación al MEB
Una de las características epidérmicas de tallos y hojas registradas con el MEB en las especies estudiadas fue la pubescencia en la superficie de la cara abaxial de hojas y tallos de E. rubrum y en toda la superficie de hojas y tallos de S. flaginoides y N. obtusifolium (Fig. 1 A-C, 2 A-C y 3 A-C).
Se observaron granos de polen de la planta madre y otros granos provenientes de árboles, arbustos y hierbas del área de estudio sobre la superficie de hojas y tallos de S. flaginoides, N. obtusifolium (Fig. 2 A-C y 3 A-C) y de M. spinosum. En este último caso la epidermis de los tallos no presenta pubescencia (Fig. 4 A-C)
Espectros polínicos de muestras de superficie de tallos y hojas
El diagrama polínico porcentual mostró como tipos polínicos dominantes, en todas las muestras de las diferentes especies analizadas, a los tipos procedentes de la planta madre. Es decir, las muestras de plantas de Empetrum rubrum, presentaron como dominante al tipo polínico E. rubrum, luego a Nothofagus, Senecio, Mulinum y Rumex.
Las muestras de las plantas de Senecio filaginoides presentaron como tipos polínicos dominantes a S. flaginoides, Mulinum spinosum y Rumex.
Las muestras de plantas de Nardophyllum obtusifolium presentaron como tipos polínicos dominantes a Mulinum y Rumex.
Por último las muestras de plantas de Mulinum spinosum presentaron como dominante al tipo polínico M. spinosum (Fig. 5).
Mulinum spinosum (zoóflo) fue el único tipo polínico presente en un porcentaje mayor al 5% en la superficie de todas las plantas analizadas. En la superficie de tallos de M. spinosum se observó, mayor abundancia de polen de esta misma especie que en la superficie de hojas y tallos de S. flaginoides, de E. rubrum y de N. obtusifolium.
Por otra parte, granos de Rumex (anemóflo) se recuperaron en alto porcentaje sobre la superficie de E. rubrum y N. obtusifolium como se mencionó previamente (Fig. 5).
En la Tabla 1 se observan los mayores valores de concentración polínica en las muestras de la planta madre. Es decir, en la superficie de tallos y hojas de Empetrum rubrum se registraron los valores más altos de concentración polínica de E. rubrum. En la superficie de tallos y hojas de Senecio flaginoides se registró el valor más alto de concentración polínica de S. flaginoides, lo mismo se observó para M. spinosum. Sin embargo, en Nardophyllum obtusifolium no se observó la mayor concentración de este tipo polínico en las muestras.
Por otro lado, se observaron valores bajos de concentración de otros tipos polínicos diferentes a los de la planta madre. Entre los anemóflos Podocarpus y Poaceae, y entre los
| Tabla 1. Concentraciones de los tipos polínicos, ordenados según tipos biológicos, identificados en las muestras de superficie de plantas de Empetrum rubrum, Senecio filaginoides, Nardophyllum obtusifolium y Mulinum spinosum. En negrita la concentración de los tipos polínicos de la planta madre. | |||||||||||||||||
| Empetrum | rubrum | Senecio | filaginoides | ■ | Nardophyllum | obtusifolium | Mulinum s | pinosum | |||||||||
| Muestras | 78 | 74 | 82 | 80 | 98 | 84 | 68 | 85 | 67 | 63 | 70 | 72 | 53 | 60 | 54 | 58 | |
| Árboles | |||||||||||||||||
| Nothofagus | 288 | 100 | 76 | 43 | 0 | 13 | 74 | 102 | 0 | 374 | 155 | 191 | 24 | 40 | 0 | 214 | |
| Podocarpus | 11 | 18 | 97 | 0 | 20 | 0 | 11 | 34 | 35 | 56 | 26 | 41 | 12 | 0 | 0 | 19 | |
| Hemiparásita | |||||||||||||||||
| Miso den drum | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 224 | 0 | 55 | 48 | 0 | 0 | 0 | |
| Arbustos | |||||||||||||||||
| Empetrum rubrum | 1461 | 236 | 1288 17831 | 0 | 0 | 85 | 34 | 0 | 0 | 142 | 68 | 12 | 0 | 0 | 136 | ||
| Gaultheria | 0 | 0 | 433 | 170 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 14 | 0 | 79 | 0 | 0 | |
| Senecio | 0 | 18 | 0 | 43 | 0 | 0 | 0 | 17 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Senecio filaginoides | 133 | 89 | 476 | 0 | 5943 | 4017 | 9985 | 2283 | 0 | 299 | 52 | 41 | 72 | 79 | 313 | 117 | |
| Baccharis | 0 | 0 | 0 | 0 | 10 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Nardophyllum obtusifolium | 0 | 0 | 11 | 0 | 0 | 0 | 0 | 0 | 0 | 2896 | 142 | 205 | 12 | 0 | 0 | 0 | |
| Nassauvia | 66 | 65 | 0 | 0 | 0 | 13 | 117 | 0 | 0 | 0 | 0 | 27 | 12 | 0 | 0 | 0 | |
| Perezia | 0 | 6 | 11 | 0 | 0 | 0 | 0 | 0 | 17 | 0 | 39 | 0 | 0 | 0 | 0 | 19 | |
| Mulinum spinosum | 144 | 207 | 43 | 0 | 341 | 225 | 1868 | 981 | 17 | 822 | 155 | 682 | 2599 | 5853 | 129904 | 22526 | |
| Azorella | 0 | 6 | 54 | 0 | 0 | 0 | 0 | 17 | 0 | 112 | 26 | 0 | 48 | 0 | 0 | 58 | |
| Acaena magellanica | 0 | 30 | 0 | 0 | 0 | 13 | 42 | 0 | 0 | 168 | 26 | 0 | 12 | 60 | 0 | 97 | |
| Hierbas | |||||||||||||||||
| Poaceae | 0 | 77 | 33 | 0 | 0 | 0 | 21 | 0 | 0 | 168 | 26 | 246 | 60 | 40 | 104 | 97 | |
| Campanulaceae | 0 | 0 | 0 | 0 | 0 | 25 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Caryophyllaceae | 0 | 0 | 0 | 0 | 0 | 25 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Cerastium | 0 | 30 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 26 | 14 | 24 | 60 | 0 | 194 | |
| Rumex | 332 | 1731 | 97 | 43 | 1004 | 388 | 891 | 17 | 105 | 1215 | 763 | 1324 | 241 | 318 | 938 | 1399 | |
| Indeterminados | 0 | 0 | 0 | 128 | 0 | 0 | 21 | 0 | 0 | 0 | 0 | 27 | 0 | 0 | 0 | 0 | |
| Concentración total | 2435 | 2611 | 2619 18257 | 7317 | 4718 | 13115 | 3484 | 174 | 6335 | 1578 | 2935 | 3177 | 6528 | 131258 | 24877 | ||

Fig. 1. Fotografías obtenidas a partir del microscopio electrónico de barrido (MEB). A: tallos y hojas de Empetrum rubrum; B: epidermis de hoja de E. rubrum; C: pelos epidérmicos del tallo de E. rubrum. Escalas= A: 1 mm; B: 100 µm; C: 50 µm.
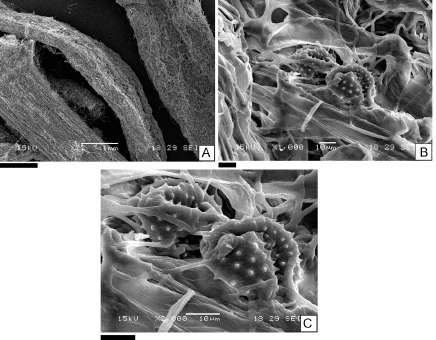
Fig. 2. Fotografías obtenidas a partir del microscopio electrónico de barrido (MEB). A: tallo y hojas de Senecio flaginoides; B: epidermis de hoja de S. flaginoides; C: granos de polen S. flaginoides. Escalas= A: 1 mm; B: 10 µm; C: 10 µm.

Fig. 3. Fotografías obtenidas a partir del microscopio electrónico de barrido (MEB). A: tallo y hojas de Nardophyllum obtusifolium; B: epidermis de hoja de N. obtusifolium; C: grano de polen de N. obtusifolium. Escalas= A: 1 mm; B: 50 µm; C: 10 µm.
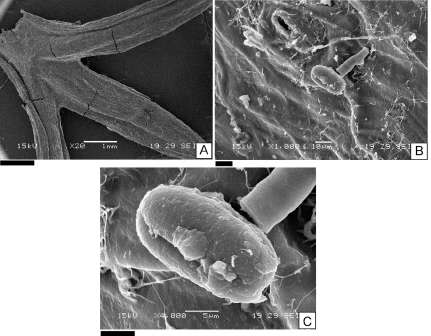
Fig. 4. Fotografías obtenidas a partir del microscopio electrónico de barrido (MEB). A: tallo de Mulinum spinosum; B: epidermis de tallo de M. spinosum; C: grano de polen de M. spinosum. Escalas= A: 1 mm; B: 10 µm; C: 5 µm.
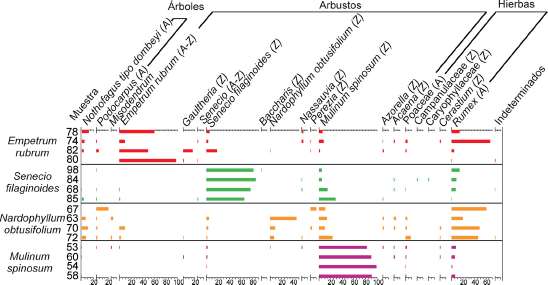
Fig. 5. Diagrama polínico porcentual de la superficie de tallos y hojas de las especies que forman parte de la dieta de Lama guanicoe en los alrededores del Cerro Casa de Piedra. (A) tipos polínicos anemóflos, (Z) zoóflos.
zoófilos Gaultheria, Nassauvia, Baccharis, Perezia, Azorella, Acaena, Caryophyllaceae y Cerastium. Las plantas de N. obtusifolium y E. rubrum presentaron mayor abundancia de polen de Nothofagus que las plantas de Senecio flaginoides y Mulinum spinosum (Fig. 5; Tabla 1).
discusion
La presencia de granos de polen de árboles, arbustos y hierbas, sobre la superficie de plantas de S. flaginoides y N. obtusifolium se relaciona con la pubescencia de la epidermis de las hojas y tallos de estas especies. Por el contrario, las plantas de Mulinum spinosum no presentan pubescencia en la superficie de sus tallos, sin embargo esta especie fue la que mayor concentración polínica presentó superando al resto de las especies.
Estos resultados concuerdan con los obtenidos por O'Rourke (1991) que menciona que diferentes especies del desierto "atrapan" polen del aire de acuerdo con las características epidérmicas de cada una de las especies, en este caso los pelos epidérmicos actuarían como trampa de granos de polen y esporas.
Con respecto a la relación polen - planta madre, la gran abundancia del tipo polínico M. spinosum sobre la superficie de la planta madre, a diferencia de lo que ocurrió en N. obtusifolium y S. flaginoides, se debería a que su pico de foración es en verano (Arroyo Kalin et al., 1981; Damascos et al., 2008), coincidente con el momento de recolección de las plantas para el estudio. M. spinosum es un subarbusto glauco, depreso-globoso, muy ramificado. Las hojas son alternas, no presentan pubescencia, son vaginantes en la base, y terminadas en 3 o 5 segmentos punzantes. Presentan fores unisexuales amarillas que sobresalen por encima del follaje (Dimitri, 1974). Tiene polinización entomófila (Forcone & Muñoz, 2009). Además, M. spinosum crece acompañado por S. flaginoides (Roig, 1998) y, por lo tanto, el polen de M. spinosum puede también quedar atrapado en los pelos de la superficie de hojas y tallos de S. flaginoides.
La alta concentración del tipo polínico Senecio sobre la superficie de S. flaginoides se debería a que esta especie forece en verano y además tiene una alta producción polínica. Además, las equinas presentes en la exina de los granos de polen le permiten adherirse con mayor facilidad a la superficie de las plantas. S. flaginoides es un arbusto hemisférico, ramoso, densamente tomentoso, con tallos hojosos hasta el ápice. Presenta hojas alternas, sésiles, con láminas lineares densamente tomentosas en ambas caras (Freire, 2014). De acuerdo a las características epidérmicas, esta especie actuaría como trampa de los granos de polen lo que explicaría la alta concentración polínica de Senecio sobre la planta madre.
Por otro lado, la baja cantidad de polen de N. obtusifolium en todas las muestras analizadas podría deberse a que esta especie forece de noviembre a enero, no coincidente con la fecha de recolección de las plantas. Probablemente el polen depositado sobre las hojas de la planta madre y de las otras especies haya sido removido por las corrientes de aire o precipitaciones. N. obtusifolium es un arbusto densamente ramoso y con abundantes hojas, las ramas viejas sin pilosidad y las jóvenes muy tomentosas. Las hojas son pequeñas y lineales. La inforescencia es un capítulo y se encuentra en el ápice de las ramas (Dimitri, 1974).
Por último, la alta concentración polínica de Empetrum rubrum sobre la superficie de las hojas y tallos de la planta madre estaría relacionada con las características epidérmicas de las plantas y la morfología de los arbustos. E. rubrum es un arbusto muy achaparrado y extendido horizontalmente con las ramas pegadas al suelo, que forma matas y posee ramitas con tomento blanquecino denso. Las hojas alternas o casi verticiladas, encontrándose muy amontonadas, cubren las ramitas que son gruesas y coriáceas. Las flores son pequeñas solitarias subsésiles y presentan estambres muy exertos (Dimitri, 1974). De acuerdo a su morfología esta especie podría actuar de trampa polínica, como se comportan los musgos.
La mayor cantidad de granos de Nothofagus sp. registrada sobre la superficie de las plantas de N. obtusifolium y E. rubrum que en las otras especies estudiadas, podría relacionarse con la presencia de árboles adultos y renovales dispersos de Nothofagus pumilio en el área de muestreo. De esta manera, el polen anemóflo de Nothofagus, con alta capacidad de dispersión, se depositaría sobre la superficie de las especies analizadas.
Por otro lado, el tipo polínico Podocarpus, podría corresponder a P. nubigena Lindl que tiene una distribución en la Cordillera de los Andes desde los 40°S hacia el sur (Donoso Zegers, 2006). La presencia de este tipo polínico en la superficie de las plantas analizadas podría deberse a que su dispersión es anemófla y llegaría hasta el área gracias a los vientos del oeste.
Por otro lado, en todas las muestras se encontró el tipo polínico Rumex que podría corresponder a Rumex acetosella de la Familia Polygonaceae que crece en el área y forece en verano.
Los valores bajos de polen de Poaceae sobre la superficie de todas las especies analizadas podría deberse a que plantas de esta familia no se encontraban en su período de polinización. El alto porcentaje y concentración de polen de Rumex en las muestras de E. rubrum y Nardophyllum obtusifolium probablemente esté asociado a la estación de polinización, ya que forece en verano.
Como síntesis se puede afrmar que los espectros polínicos de la superficie de algunas plantas, que son parte de la dieta de las poblaciones de guanaco del PNPM, mostraron que los tipos polínicos dominantes proceden de la planta madre, salvo en las muestras de N. obtusifolium, donde M. spinosum dominó el espectro polínico. Además, en todas las muestras fue importante la abundancia de otros tipos polínicos tanto con dispersión anemófla o zoófla.
Estos resultados muestran la magnitud del aporte de granos de polen, a partir del consumo de órganos vegetativos de diferentes ítems alimentarios, a las heces de los guanacos y de esta manera brindan información para la interpretación del espectro polínico de heces y coprolitos de guanaco para estudios de dieta y paleodieta que se están llevando a cabo en Patagonia (Velázquez, 2016; Velázquez et al., 2017). Asimismo este análisis permite conocer la composición del espectro polínico que los guanacos pudieron haber ingerido accidentalmente por estar depositado sobre la superficie de los ítems alimentarios. Una de las hipótesis que surge es que si los guanacos se alimentan de plantas de Empetrum rubrum, Senecio flaginoides y Mulinum spinosum, cuyo polen puede quedar atrapado entre los pelos epidérmicos de las hojas y tallos, probablemente una alta concentración polínica de estas especies en las heces estaría sugiriendo que estas especies formaron parte de la dieta del guanaco. Distinto es el caso de N. obtusifolium, que presentó un mayor porcentaje y concentración de Mulinum en sus hojas y tallos que de granos procedentes de la planta madre. Sería necesario el análisis de un mayor número de muestras de cada una de las especies analizadas, recolectadas en diferentes estaciones del año, para evaluar el efecto de la producción y la estación de polinización sobre la abundancia de granos de polen que quedan atrapados en la superficie de las hojas y tallos. Además sería necesario evaluar cuánto tiempo permanecen en la superficie de las plantas hasta que el viento, turbulencias o las precipitaciones los remuevan.
conclusiones
El análisis de la contaminación polínica de algunos ítems alimenticios de los guanacos del PNPM permitió identificar sesgos en las asociaciones polínicas que podrían enmascarar la determinación de los ítems consumidos intencionalmente y no de forma accidental.
Los espectros polínicos de la superficie de plantas que son parte de la dieta de guanacos del PNPM mostraron que los tipos polínicos dominantes en todas las muestras proceden de la planta madre, salvo el caso de Nardophyllum obtusifollium.
Asimismo, la recuperación de polen anemóflo y zoóflo de especies diferentes a la planta madre, aunque en baja concentración, sobre la superficie de hojas y tallos pubescentes de especies que forman parte de la dieta del guanaco permitió conocer los tipos polínicos que podrían haber sido ingeridos accidentalmente. Estos resultados mostraron la abundancia y diversidad de polen proveniente de la lluvia polínica sobre el alimento. Estos espectros polínicos están relacionados a la estación de polinización y a la cercanía de las plantas. La contaminación del alimento con polen anemóflo arbóreo podría deberse a la alta capacidad de dispersión del polen como también a la arquitectura de las especies que actúan como trampas de polen. Por otro lado, la alta concentración de polen (tanto zoóflo como anemóflo) sobre la superficie de la planta madre está relacionada a sus características epidérmicas, como la presencia de pelos que actúan como trampas polínicas.
Resta ampliar el número de muestras para poder evaluar los efectos de a) la producción polínica y b) el tiempo de permanencia de los granos en la superficie de las plantas, sobre la abundancia de granos que quedan atrapados en la superficie de los órganos vegetativos de las diferentes especies que son fuentes alimenticias de los guanacos.
agradecimientos
Al Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET); Universidad Nacional de Mar del Plata (UNMDP), Proyecto (EXA-UNMDP EXA 855/18); Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT), Fondo para la Investigación Científica y Tecnológica (FONCyT), Proyecto (PICT No. 2015 2541, L.Caruso Fermé; PICT No. 2017 3239, N.J. Velázquez; PICT No. 2017 0455, L.S. Burry). A Belén Martel quien colaboró con el procesamiento de las muestras, identificación y recuento polínico durante una pasantía de investigación en el Laboratorio de Palinología y Bioantropología, bajo la dirección de Lidia Susana Burry y la tutoría de Nadia Jimena Velázquez. A los revisores anónimos que con sus sugerencias permitieron mejorar el manuscrito.
Bibliografia
ALCOVER, J. A. R., E. Yll. PEREZ-OBIOL & P. BOVER. 1999. The diet of Myotragus balearicus Bate 1909 (Artiodactyla: Caprinae), an extinct bovid from the Balearic Islands: evidence from coprolites. Biol. J. Linn. Soc. Lond. 1: 57-74. https://doi.org/10.1111/j.1095-8312.1999.tb01917.x [ Links ]
ARROYO KALIN, M. T., J. ARMESTO & C. VILLAGRAN. 1981. Plant phenological patterns in the High Andean Cordillera of Central Chile. J. Ecol. 69: 205-223. [ Links ]
BANK, M. S., R.J. SARNO, N. K. CAMPBELL & W. L. FRANKLIN. 2002. Predation of guanacos (Lama guanicoe) by southernmost mountain lions (Puma concolor) during a historically severe winter in Torres del Paine National Park, Chile. J. Zool. 258: 215-222. https://doi.org/10.1017/S0952836902001334 [ Links ]
BRYANT Jr., V. M. & R. G., HOLLOWAY. 1983. The role of palynology in archaeology. In: CHAVES, S.A.M. & K.J. REINHARD, 2006. Critical analysis of coprolite evidence of medicinal plant use, Piauí, Brazil. Palaeogeogr. Palaeoclimatol. Palaeoecol. 237: 110-118. https://doi.org/10.1016/B978-0-12-003106-1.50010-9 [ Links ]
BURGI, M. V. 2007. Radio de acción del guanaco (Lama guanicoe) en el NE de Chubut. Mastozool. neotrop. 14 (2): 285-291. [ Links ]
CARRIÓN, J. S., G. GIL, E. RODRÍGUEZ, N. FUENTES, M. GARCÍA-ANTÓN & A. ARRIBAS. 2005. Palynology of badger coprolites from central Spain. Palaeogeogr. Palaeoclimatol. Palaeoecol. 226: 259-271. https://doi.org/10.1016/j.palaeo.2005.05.016 [ Links ]
CARRIÓN, J. S., J. A., RIQUELME, C. NAVARRO & M. MUNUERA. 2001. Pollen in hyaena coprolites refects late glacial landscape in southern Spain. Palaeogeogr. Palaeoclimatol. Palaeoecol. 176: 193-205. https://doi.org/10.1016/S0031-0182(01)00338-8 [ Links ]
CARRIÓN, J. S., R. YII, J. A. RIQUELME & P. GONZÁLEZ. 2004. Perspectivas del análisis polínico de coprolitos y otros depósitos biogénicos útiles en la inferencia paleoambiental. Miscelánea en Homenaje a Emiliano Aguirre: Paleontología. Museo Arqueológico Regional, Madrid, pp. 128-139.
CHAVES, S. A. M. & K. J. REINHARD (eds.). 2006. Critical analysis of coprolite evidence of medicinal plant use, Piauí, Brazil. Palaeogeogr. Palaeoclimatol. Palaeoecol. 237: 110-118. https://doi.org/10.1016/j.palaeo.2005.11.031
CHAVES, S. A. M. 2000. Estudo palinológico de Coprólitos pré-históricos Holocenos coletados na toca do Boqueirão do sítio da Pedra Furada-contribuições paleoetnológicas, paleoclimáticas e paleoambientais para a região sudeste do Piauí-Brasil. Revista do Museu de Arqueología e Etnología, Universidade de São Paulo 10: 103-120.
CORREA, M. V. (dir.). 1971-1999. Flora Patagónica. Colecc. Ci. INTA, Buenos Aires.
DAMASCOS, M. A., D. BARTHELEMY, C. EZCURRA, P. MARTÍNEZ & C. BRION. 2008. Plant phenology, shoot growth, and branching pattern in Mulinum spinosum (Apiaceae), a cushion shrub of the arid Patagonian steppe of Argentina. J. Arid Environ. 72: 1977-1988. https://doi.org/10.1016/j.jaridenv.2008.07.001
D'ANTONI, H. L. 1979. Arqueoecología: El hombre en los ecosistemas del pasado a través de la Palinología. Colección Científica de Arqueoecología. México. 134 pp.
D'ANTONI, H. 2008. Arqueoecología: Sistémica y Caótica. Consejo Superior de Investigaciones Científicas (CSIC), Madrid.
DE KLERK, P. & H. JOOSTEN. 2007. The difference between pollen types and plant taxa: a plea for clarity and scientific freedom. Eiszeitalter und Gegenwart / Quaternary Science Journal. 56: 162-171. http://dx.doi.org/10.23689/fdgeo-1295
DIMITRI, J. M. 1974. Pequeña fora ilustrada de los Parques Nacionales andino-patagónicos, Anales de Parques Nacionales 13: 1-122.
DIMITRI, M. 1987. Enciclopedia Argentina de Agricultura y Jardinería. Edit. Acme, Buenos Aires.
DONOSO ZEGERS, C. 2006. Las Especies arbóreas de los Bosques Templados de Chile y Argentina. Autoecología. Marisa Cúneo Ediciones, Valdivia, Chile.
FAEGRI, K. & J. IVERSEN. 1989. Textbook of Pollen Analysis 4th Edition, En: FAEGRI, K., KALAND, P.E., & K., KRZYWINSKI (eds.). John Wiley and Sons, Chichester, 328 pp.
FERNÁNDEZ-JALVO, Y., L. SCOTT, J. S. CARRIÓN, G., GIL-ROMERA, J. BRINK, F. NEUMANN & L. ROSSOUW. 2010. Pollen taphonomy of hyaena coprolites: an experimental approach. First Meeting on Hyaena Dens in archaeological sites of the Iberian Peninsula. Museo Arqueológico Regional, Alcalá de Henares, pp. 149-156. https://doi.org/10.1016/S0031-0182(01)00338-8
FORCONE, A. & M. MUÑOZ. 2009. Floración de las especies de interés apícola en el noroeste de Santa Cruz, Argentina. Bol. Soc. Argent. Bot. 44: 3-4.
FRANKLIN, W. L. 1982. Biology, ecology, and relationship to man of the South American camelids. En: MARES, H. & M.G. GENOWAYS (eds.), Mammalian Biology in South America. University of Pittsburgh. Special Publication Series, 6: 457-488.
FRANKLIN, W. L. 1983. Contrasting socioecologies of South America's wild camelids: The vicuña and guanaco. En: EISEMBERG, S. F. & D. G. KLEIMAN, (eds.), Advances in the Study of Mammalian Behaviour, Special Publication, 7, pp. 573-629.
FREIRE, S. E. 2014. Flora Vascular de la República Argentina 7(3): Dicotyledoneae-Asteraceae (Senecioneae a Vernonieae). ZULOAGA, F. O., BELGRANO M. J. & A.M. ANTON (eds.) Instituto de Botánica Darwinion. http://www.jstor.org/ stable/j.ctt16xwb7t.
HEUSSER, C. J. 1971. Pollen and Spores from Chile. Modern Types of Pteridophyta, Gymnospermae and Angiospermae. University of Arizona Press, Tucson. 167 pp.
HORROCKS, M., G. J. IRWIN, M. S. MCGLONE, S. L. NICHOL & L. J. WILLIAMS. 2003. Pollen, Phytoliths and diatoms in Prehistoric Coprolites from Kohika, Bay of Plenty, New Zealand. J. Archaeol. Sci. 30: 13-20. https://doi.org/10.1006/jasc.2001.0714
HORROCKS, M., J. SALTER, J. BRAGGINS, S. NICHOL, R. MOORHOUSE & G., ELLIOTT. 2008. Plant microfossil analysis of coprolites of the critically endangered kakapo (Strigops habroptilus) parrot from New Zealand. Rev. Palaeobot. Palynol. 149: 229-245. https://doi.org/10.1016/j.revpalbo.2007.12.009
JOOSTEN, H. & P. DE KLERK. 2002. What's in a name? Some thoughts on pollen classification, identification, and nomenclature in Quaternary palynology. Rev. Palaeob. Palyn. 122: 29-45. https://doi.org/10.1016/S0034-6667(02)00090-8
LAUENROTH, W. K. 1998. Guanacos, spiny shrubs, and the evolutionary history of grazing in the Patagonian steppe. Ecología Austral 8 (2): 211-215.
MARKGRAF, V. & H. L. D'ANTONI. 1978. Pollen Flora of Argentina. Modern Spore and Pollen Types of Pteridophyta, Gymnospermae and Angiospermae. The University of Arizona Press, Tucson, 208 pp.
MOE, D. 1983. Palynology of sheep's faeces: relationship between pollen content, diet and local pollen rain. Grana 22 (2): 105-113. https://doi.org/10.1080/00173138309431970
MOORE, P. D., J. A. WEBB & M. E. COLLINSON. 1991. Pollen Analysis, second ed. Blackwell, London.
MUÑOZ, A. 2008. Guanaco (Lama guanicoe) browsing on lenga (Nothofagus pumilio) regeneration in Tierra del Fuego. Tesis para optar el título de Magíster en Ciencias Biológicas, con mención en Ecología y Biología Evolutiva. Universidad de Chile. 37pp.
O' ROURKE, M. K. 1991. Pollen in packrat middens. Grana 30 (2): 337-341. https://doi.org/10.1080/00173139109431988
PARDIÑAS, U. F. J. 1998. Roedores holocénicos del sitio Casa de Piedra 5 (Santa Cruz, Argentina): tafonomía y paleoambientes. Palimpsesto. Revista de Arqueología 5: 66-90.
PUIG, S., F. VIDELA, S. MONGE & V. ROIG.1996. Seasonal variations in guanaco diet (Lama guanicoe Müller 1776) and food availability in Northern Patagonia, Argentina. J. Arid Environ. 34: 215-224. https://doi.org/10.1006/jare.1996.0103
RAEDEKE, K. J. 1978. El guanaco de Magallanes, Chile, su distribución y biología Corporación Nacional Forestal, Publicación Técnica Nº 4, Departamento
de Conservación del Medio Ambiente, Santiago, Chile. 182 pp.
RAEDEKE, K. J. 1980. Food habits of the guanaco (Lama guanicoe) of Tierra del Fuego, Chile. Turrialba 30: 177-181.
REINHARD, K. J. & V. M. J. BRYANT. 1992. Coprolite analysis: a biological perspective on archaeology, In: SHIFFER, M. (eds.), Advances in Archaeological Method and Theory 4. University of Arizona Press. Tucson, pp. 245-288.
ROIG, F. A. 1998. La vegetación de la Patagonia. En: Correa M (compaginador), Flora Patagónica. Colección Científica INTA. Tomo VIII (1): 48-174.
TAUBER, H. 1967. Investigations of the mode of pollen transfer in forested areas. Rev. Palaeob. Palyn. 3: 277-286.
VELÁZQUEZ, N. J. & L. S. BURRY. 2012. Palynological analysis of Lama guanicoe modern feces and its importance for the study of coprolites from Patagonia, Argentina. Rev. Palaeob. Palyn. 184: 14-23. https://doi.org/10.1016/j.revpalbo.2012.07.012
VELÁZQUEZ, N. J., L. S. BURRY, M. V. MANCINI, & M. H. FUGASSA. 2010. Coprolitos de camélidos del Holoceno como indicadores paleoambientales. Magallania 38: 213-229. http://dx.doi.org/10.4067/S0718-22442010000200013
WHEELER, J. C. 1991. Origen, evolución y status actual. En: FERNÁNDEZ-BACA, S., (ed.), Avances y perspectivas del conocimiento de los camélidos sudamericanos: 11-48. Oficina Regional de la FAO para América Latina y el Caribe, Santiago, Chile.
WOOD, J. R., J. M. WILMSHURST, S. J. WAGSTAFF, T. H. WORTHY, N. J. RAWLENCE & A. COOPER. 2012. High-Resolution Coproecology: Using Coprolites to Reconstruct the Habits and Habitats of New Zealand's Extinct Upland Moa (Megalapteryx didinus). PLoS ONE 7: 1-13. https://doi.org/10.1371/journal.pone.0040025

