










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkBoletín de la Sociedad Argentina de Botánica
versão On-line ISSN 1851-2372
Bol. Soc. Argent. Bot. vol.57 no.3 Córdoba set. 2022
http://dx.doi.org/10.31055/1851.2372.v57.n3.37652
10.31055/1851.2372.v57.n3.37652
Articulos
Las plantas que alimentan el fogón: primeros resultados del registro fitolítico de estructuras de combustión de Cueva de los Catalanes (Araucanía, Chile)
Plants that feed the hearth: first results of the phytolith record of combustión structures from Cueva de los Catalanes (Araucanía, Chile)
Constanza Roa Solís 1
Débora Zurro 2
1. División de Arqueología Museo de La Plata (Laboratorio Arqueobotánica), Facultad de Ciencias Naturales y Museo, Universidad Nacional de la Plata, Av. 122 y Av. 60, La Plata, Argentina.
2. HUMANE-Human Ecology and Archaeology Research Group. Institució Mila i Fontanals de Investigación en Humanidades, Consejo Superior de Investigaciones Científicas (IMF-CSIC), Carrer Egipcíaques, 15, Barcelona, España.
*c.roasolis@gmx.com
Summary
Background and aims: Fire management is a widely discussed topic in Archaeology, which has been mainly addressed through anthracology and carpology. Phytolith analysis complements these techniques thanks to some silica phytolith advantages: 1) they are preserved in a wide variety of environments without requiring carbonization or special conditions and 2) they account for different taxonomic categories and different plant organs. The present study seeks to identify the plant specific signal from the combustion structures of Cueva de los Catalanes, in comparison to areas without evidence of combustion and throughout the occupational sequence.
M&M: The material analyzed corresponds to sediment samples from Cueva de Los Catalanes's different occupation levels and combustion structures. Phytolith analysis and the statistical processing of the data were carried out.
Results: This work suggests that phytolith assemblages are very heterogeneous. However, samples coming from the combustion structures show both higher concentrations and diversity of phytoliths. Likewise, pre-Hispanic samples are richer in phytoliths and the spectrum shows some different features, which correlate with higher densities of the archaeological materials.
Conclusions: Phytolith analysis is outlined as a useful as well as a complementary tool to understand the study area's fire management, having a wide potential poorly explored.
Key words: Archaeobotany, Ceramic Period, fire management, hearth, Late Holocene, phytoliths.
Resumen
Introducción y objetivos: El manejo del fuego es una temática ampliamente tratada en Arqueología, generalmente mediante análisis antracológicos y carpológicos. El análisis de fitolitos complementa a estas técnicas gracias a ventajas de los silicoficolitos: 1) se conservan en muy variados ambientes, sin requerir carbonización ni condiciones especiales; 2) dan cuenta de distintas categorías taxonómicas y de distintos órganos de las plantas. El presente estudio busca identificar la señal vegetal específica de las estructuras de combustión de la Cueva de los Catalanes, en comparación a áreas sin evidencias de combustión y a lo largo de la secuencia ocupacional.
M&M: El material analizado corresponde a muestras de sedimento de los distintos niveles de ocupación y de las estructuras de combustión de Cueva de los Catalanes. Se llevó a cabo el análisis de fitolitos y el procesamiento estadístico de los datos.
Resultados: Se sugiere que los conjuntos fitolíticos se comportan de manera muy heterogénea. No obstante, aquellos provenientes de las estructuras de combustión presentan mayores abundancias y diversidad de silicofitolitos. Asimismo, las muestras prehispánicas son más ricas en fitolitos y el espectro muestra algunas características diferentes, que se correlacionan con mayores densidades de los materiales arqueológicos.
Conclusiones: El análisis de silicofitolitos se presenta como una herramienta útil y complementaria para entender el manejo del fuego en la región de estudio, con amplio potencial pobremente explorado.
Palabras clave: Arqueobotánica, fitolitos, fogón, Holoceno Tardío, manejo del fuego, Período Alfarero.
Introducción
La presencia de hogares ha sido tradicionalmente el principal criterio diagnóstico que ha permitido identificar un contexto como escenario de una ocupación humana. El fuego forma parte intrínseca de la historia de nuestra especie (Barbetti, 1986) y es prácticamente una constante en contextos arqueológicos muy diferentes, como herramienta que ha servido al ser humano en distintos propósitos (Oakley, 1955; Perles, 1977; Pérez de Micou, 1991; Galanidou, 2000; Lumley, 2006; Frank, 2012; Massone, 2017). Así, el estudio tanto de las estructuras de combustión como de las especies y materias usadas como combustible son temas ampliamente tratados a lo largo de la historia de la Arqueología (Leroi-Gourhan & Brézillon, 1972; March, 1995; Théry-Parissot, 1998; March & Lucquin, 2007; Vallverdú et al., 2012).
Una estructura de combustión (Pérez de Micou, 1991; Frank, 2012) comprende una base de sedimentos termo-alterados y una parte superior con cenizas y carbones (Albert et al., 2000; Berna et al., 2007; Cabanes et al., 2007). El contenido de una estructura reúne tanto el material involucrado en la combustión, que proviene directamente de la preparación y alimentación del fuego, y el material derivado de las actividades desarrolladas en ella, pues estos espacios suelen ser utilizados como lugares de descarte dentro de un asentamiento (Binford, 1991 [1983]; Hansen, 2001), siendo un resumen (o fotografía) de todo tipo de materiales empleados en la ocupación. Ahora bien, durante el acondicionamiento, el encendido y el mantenimiento del fuego, se utilizan principalmente productos de origen vegetal (salvo contadas excepciones o contextos muy específicos). Estos productos vegetales podrán corresponder a plantas leñosas, y también a tallos y hojas de plantas herbáceas, que generalmente no generan residuos macrobotánicos carbonizados.
Dentro de los microrestos botánicos se encuentran los silicofitolitos: réplicas mineralizadas de células vegetales. Estos restos permanecerán en los sedimentos en muy variables contextos de depositación, ya sea como resultado de actividades antrópicas o fenómenos naturales, sin ser alterados por el decaimiento orgánico (Osterrieth et al., 2009; Zurro, 2011; Madella & Lancelotti, 2012).
En este sentido, el análisis de silicofitolitos presenta una ventaja específica que lo diferencia de otras técnicas definidas por el estudio de órganos y tejidos específicos como la Carpología o la Antracología, ya que permiten caracterizar la variabilidad del registro vegetal tanto taxonómica como anatómicamente (raíces, tallos, hojas, flores, frutos) (Metcalfe, 1969; Twiss et al., 1969; Bozarth, 1992; Ollendorf, 1992; Piperno & Pearsall, 1993; Kealhofer & Piperno, 1998; Piperno & Stothert, 2003; Ball et al., 2006; Piperno, 2006; Ball et al., 2009). De esta manera, los conjuntos fitolíticos tanto del material carbonizado como de las cenizas permiten caracterizar distintas partes y tipos de plantas que alimentaron la estructura de combustión en el desarrollo de actividades específicas. Esto es relevante por cuanto sabemos que los hogares no siempre cumplen la misma función, ya que pueden estar dirigidos a la producción de calor, de luz, cocción, ahumado de alimentos o generación de señales de humo, de manera que los combustibles pueden variar en función de estas elecciones y las características de los combustibles disponibles (Lejay et al., 2016). Por tanto, para el estudio del material vegetal combustionado, los silicofitolitos representan una excelente línea de evidencia, que contribuye a un estudio arqueobotánico integral del manejo del fuego.
En Suramérica, específicamente en la Patagonia, se han estudiado las estructuras de combustión en contextos cazadores-recolectores desde distintas perspectivas etnoarqueológicas, contextuales y experimentales (March et al., 1989; March et al., 1991; Pérez de Micou, 1991; March, 1992; Massone et al., 1998; Frank, 2012; Massone, 2017; Morales et al, 2017; Hammond & Ciampagna, 2019). En cuanto a los residuos vegetales, se han estudiado principalmente desde la antracología (Solari, 1991, 1992, 1993, 1994, 2000, 2003, 2009; Solari et al, 2002; Solari & Lehnebach, 2004; Rojas, 2004; Zurro et al., 2009; Caruso Fermé 2013a, 2013b; Caruso Fermé & Civalero 2014, 2019; Ciampagna et al, 2016; Massone & Solari, 2017; Caruso Fermé & Zangrando, 2019).
La mayor parte de los grupos que habitaron históricamente el gran territorio de Patagonia detentaban un modo de vida cazador-recolector, o bien pescador-cazador-recolector, y generaron registros materiales específicos en relación a un modo de vida de amplia movilidad. No obstante, hacia el Holoceno tardío la parte septentrional de este territorio (o Norpatagonia) conformó el Wallmapu o "País Mapuche", siendo habitado por este pueblo a ambos lados de la cordillera de los Andes (Argentina y Chile). En esta zona se inserta el área sur de Chile, donde durante los últimos dos mil años habitaron poblaciones con modos de vida hortícola a agrícola, presentando mayor nucleamiento poblacional (Quiroz & Sánchez, 1997; Adán & Mera, 2011; Campbell, 2014; Campbell et al., 2018a). Las particularidades de estas poblaciones, suponemos, han incidido en los registros de estructuras de combustión arqueológicas. Dichas estructuras son espacios dentro del sitio arqueológico que corresponden, además, a lugares de agregación social. En el área mapuche, en específico, se reconoce la importancia del fogón como área de socialización, usada para calentarse, iluminarse y cocinar, siendo además un espacio manejado por mujeres (Egert & Godoy, 2008).
Como discutimos más profundamente en otros trabajos (Roa, 2018; Roa & Capparelli, inéd.), el estudio de las plantas involucradas en el manejo del fuego y, con esto, el estudio del uso de plantas y la relación de las comunidades con su entorno, presenta grandes potencialidades en el sur de Chile, especialmente en la región de la Araucanía gracias al conocimiento etnobotánico de que disponemos. En esta área ha habitado históricamente el pueblo mapuche, el cual se ha relacionado con su entorno no-humano de manera respetuosa, llevando a cabo la recolección de plantas en conjunto con su cultivo, lo que además se ve complementado con la gran biodiversidad del bosque templado. Se recolectaron plantas para diversos fines, incluidas distintas etapas del manejo del fuego, utilizándose plantas herbáceas y leñosas como combustibles e iniciadores (Roa & Capparelli, inéd.). Sin embargo, la relación entre el ser humano y el paisaje ha cambiado de manera drástica en los últimos dos siglos con el advenimiento de los colonos chilenos y europeos en el territorio (Solari et al, inéd.); en la actualidad, esta zona se encuentra invadida por la industria forestal, la que incentiva la plantación de especies arbóreas de rápido crecimiento, que requieren de mucha agua y acidifican la tierra de manera acelerada, en detrimento del bosque nativo y de las reservas de agua. Esto ha conllevado un empobrecimiento económico y social de las comunidades mapuches, cuyos espacios históricos de recolección se han visto mermados.
Este trabajo busca evaluar la potencialidad del análisis de fitolitos en estructuras de combustión de sitios arqueológicos de Araucanía. Por una parte, y desde un punto de vista metodológico, se plantea la posibilidad de identificar una señal específica en el input vegetal de estructuras de combustión, de manera cualitativa y cuantitativa, que las haga distinguibles de aquellos contextos de muestreo que no corresponden a rasgos arqueológicos (y que hemos definido como "muestras blancas"). Dada la posible multifuncionalidad de los hogares, así como la posibilidad de que a los restos se sumen restos de lo cocinado o deshechos de la limpieza de los suelos de ocupación, desde un punto de vista metodológico en las estructuras de combustión se esperaría encontrar una mayor abundancia y diversidad de morfotipos de fitolitos, así como una mayor concentración de microrestos de plantas leñosas (dicotiledóneas).
En segundo lugar, y desde un punto de vista substantivo, se pretende evaluar el uso de plantas involucradas en el manejo del fuego a lo largo de la secuencia ocupacional de la cueva, atendiendo a los cambios histórico-culturales que la secuencia representa, sugeridos por otros materiales arqueológicos (por ejemplo, cambios entre los períodos Alfarero temprano y tardío, o entre los niveles pre y post-contacto). A modo de hipótesis, en base a la inexistencia de estructuras de combustión en la cueva en momentos posteriores a la colonización europea y, a la ostensible disminución en la densidad de materiales arqueológicos, suponemos que el input vegetal disminuiría en relación al período prehispánico.
Materiales y Métodos
Area de Estudio
La Cueva de los Catalanes (37° 48' lat. S; 72° 29' long. W; 130 msnm) de Lolcura Bajo (comuna de Collipulli, región de la Araucanía), se ubica en el valle central en la cuenca del río Renaico (Fig. 1A). Esta área corresponde a una zona climática transicional entre los macro-bioclimas Mediterráneo y Templado (Luebert & Pliscoff, 2017), combinando la distribución del bosque templado y elementos del bosque esclerófilo.
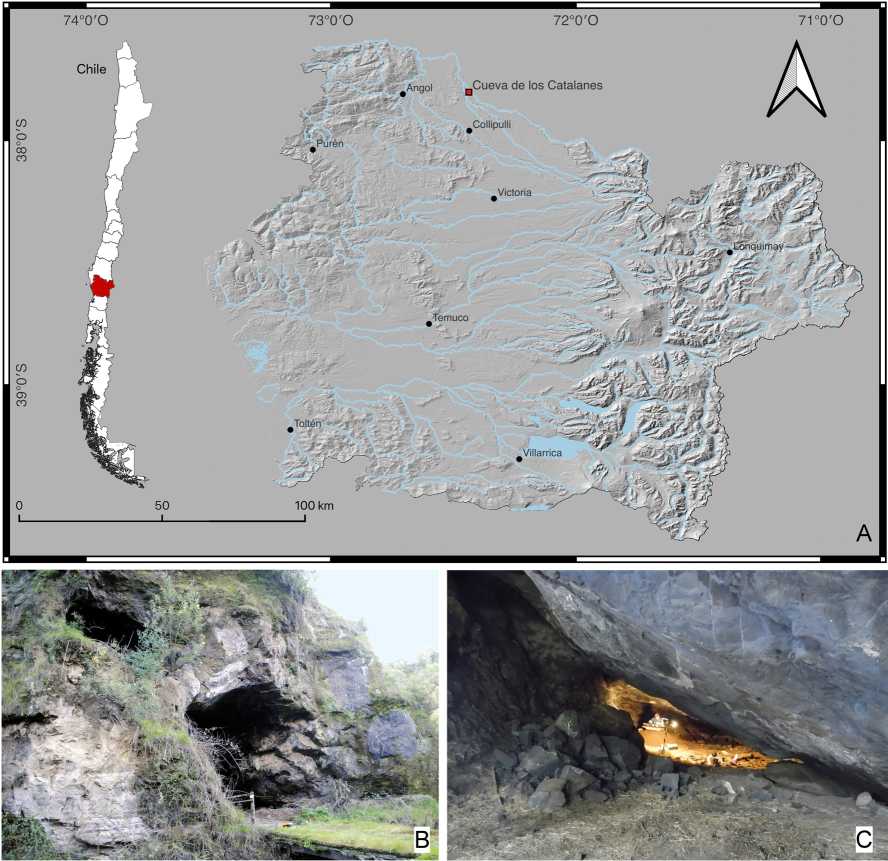
Fig. 1. Cueva de los Catalanes A: Área de emplazamiento de la cueva (Región de la Araucanía). B: Entrada de la cueva donde se observan sus dos aperturas. C: Vista del interior desde la entrada con un angostamiento que separa entrada e interior. B: foto de C. Dávila, C: foto de A. Peñaloza.
Estudios de paleoambiente en los valles de Purén y Lumaco cercanos a la cueva, describen que la composición florística se ha mantenido más o menos similar desde hace cuatro mil años (Abarzúa et al, 2014), aunque ciertos cambios se han percibido durante los últimos dos mil años en la zona cordillerana, presentando una tendencia hacia condiciones de mayor sequedad, con un período más húmedo entre 660-1561 d.C. (Torres et al., 2008). Actualmente, la comuna de Collipulli presenta importantes áreas con actividad silvícola (52,14%), además de agroganadera (29,08%), las que ocupan la mayor parte del territorio1. 1 2
Yacimiento
La cueva se extiende a lo largo de 33 m y presenta dos zonas bien diferenciadas: la entrada de 12 m de ancho y 5 m de alto, y la sección interior cuya altura promedio es de 2 m (Fig. 1B-C). Inmediatamente afuera de la cueva corre un canal de regadío construido posiblemente durante la primera mitad del siglo XX.
Este yacimiento fue excavado en dos ocasiones, la primera de ellas en 1956 (Menghin, 19591960; Berdichewsky, 1968) y la segunda en 2016 (Campbell et al, 2017). Las muestras del presente estudio fueron obtenidas a partir de esta última intervención. En 2016, se excavaron cuatro unidades de 1 x 1 m (Fig. 2A), de las cuales tres se dispusieron en un eje lineal al interior de la cueva (P1, P2, P3); la última se excavó fuera de la línea de goteo (P4), en la otra orilla del canal de regadío y resultó presentar una estratigrafía completamente removida (Campbell et al, 2018b; Roa, 2018). El grueso de la ocupación se concentra en las unidades P1 y P2 (Fig. 2B).
En base a los fechados absolutos y a la distribución de los materiales, se ha propuesto en trabajos previos una ocupación continua desde una fecha probablemente anterior a los 637-759 años cal. d.C. hasta la actualidad (Campbell et al, 2018b). En base a los resultados de la estratigrafía, fechados y materiales arqueológicos, se segregó una ocupación prehispánica que incluye las capas L10 a L5, y una ocupación post-contacto europeo o histórica entre las capas L4 a L1 (Roa, 2018). La ocupación prehispánica exhibió fechas durante el período Alfarero, presentando materiales como alfarería y líticos concordantes con las tradiciones descritas para este período, además de cultivos nativos (Zea mays L., Chenopodium quinoa Willd., Phaseoulus vulgaris L., Madia sativa Molina) comunes a otros sitios del área sur del país. Por otro lado, la ocupación post-contacto se diferencia por la aparición de cultivos europeos (Triticum sp.) y algunos elementos de cultura material introducida como loza. No obstante, esta ocupación presenta materiales de factura indígena no del todo diferenciables de los períodos precedentes (Gajardo, 2016; González, 2018). En los niveles prehispánicos, hemos diferenciado las primeras ocupaciones correspondientes al período Alfarero temprano (3501000 d.C.; capas L10 a L8.3) y las siguientes durante el período Alfarero tardío (1000-1550 d.C.; capas L8.2 a L5). Llama la atención que la mayor parte de los materiales arqueológicos provienen de los niveles del período Alfarero temprano, disminuyendo hacia el período Alfarero tardío, con un aumento aparente durante momentos post-contacto (Gajardo, 2016; González, 2018; Roa, 2018; Peñaloza, com. pers.).
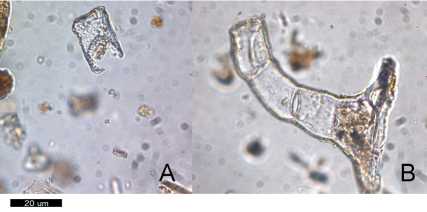
Fig. 2. Plano de excavación y crono-estratigrafía de la Cueva de los Catalanes. A: Plano de la excavación del año 2016 con las unidades de excavación (gris) en vistas lateral (arriba) y en planta (abajo). B: Resumen crono-estratigráfico y toma de muestras.
Muestreo
En el momento de la excavación las muestras no fueron tomadas para el análisis de fitolitos, por lo que no se dispone de todas las muestras que un muestreo dirigido habría proporcionado. No obstante lo anterior, el muestreo posibilita abordar nuestras preguntas ya que nuestro interés radica en identificar criterios diagnósticos para las muestras de combustión. Así, se analizaron tanto muestras procedentes de estos rasgos (en las que a priori hay una alta expectativa de obtener resultados relativos a la gestión de recursos vegetales), como aquellas procedentes de áreas carentes de rasgos específicos (muestras blancas) y que nos permitirán caracterizar de forma general el contexto en estudio. Con todo, el conjunto analizado corresponde a 31 muestras de sedimentos arqueológicos, de las cuales 20 provienen del muestreo vertical de un perfil estratigráfico (P1), nueve de estructuras de combustión (de las cuales seis son muestras de rasgos de ceniza y tres muestras de rasgos de sedimento termo-alterado; P1, P2, P3) y las dos restantes de sedimentos asociados a tubos de pipa y carporrestos (P2) (Tabla 1).
Análisis de fitolitos
El análisis de silicofitolitos se desarrolló en el Laboratorio BIOGEOPAL de la Institució Milá i Fontanals (CSIC). Desde una muestra aproximada de 4 g de sedimento se extrajeron los silicofitolitos de acuerdo al protocolo de Madella et al. (1998). Este procedimiento incluye la remoción de carbonatos con HCl, la defloculación de la muestra con calgón (NaPO3)6 y la eliminación de la materia orgánica con peróxido de hidrógeno (H2O2). Luego de estos pasos se obtiene la fracción insoluble en ácido (Acid Insoluble Fraction, en adelante AIF) (Albert & Weiner, 2001). Finalmente, se lleva a cabo la separación densimétrica de los minerales con SPT (Na6(H2W12O40)H2O), obteniendo el residuo de sílice amorfo, donde permanecen los silicofitolitos (Tabla 1).
El residuo de peso conocido (0,5-1,7 mg) fue montado en láminas con el medio fijo DPX Mountant (Sigma-Aldrich®). El conteo de silicofitolitos y tejidos se llevó a cabo hasta completar un total de 250 silicofitolitos aislados por lámina (Zurro, 2018). Este conteo se realizó según transectas dentro de la lámina, registrando el porcentaje revisado de la lámina donde se encontraba la suma de 250 silicofitolitos, para luego extrapolar el número de silicofitolitos por lámina y, por gramo de sedimento, de acuerdo al peso montado (Albert & Weiner, 2001; Zurro, 2011).
La determinación morfo-tipológica se realizó con un microscopio Leica bajo aumentos de 200x y 630x, utilizando descriptores del International Code of Phytolith Nomenclature 1.0 (Madella et al, 2005), que considera la proveniencia taxonómica y anatómica (Tabla 2). De acuerdo a su nivel de determinación, se consideraron dos categorías clasificatorias: determinado y no-determinado. En el caso de la primera, corresponde a aquellos fitolitos que fueron adscritos a categorías morfo-tipológicas concretas, con valor taxonómico, anatómico o ambos; mientras que la segunda da cuenta de los fitolitos que no se corresponden con una categoría morfo-tipológica particular, ya sea a causa de su estado de silicificación incompleta o tafonomización, como a sus morfologías variables o no-consistentes (sensu Albert et al., 2000).
Adicionalmente, se evaluó el daño tafonómico (mecánico o químico) para cada morfotipo, desdoblándolos en tafonomizados y no tafonomizados (los datos se presentan de acuerdo al porcentaje de tafonomización del conjunto) (Madella & Lancelotti, 2012).
Tabla 1. Descripción de las muestras y resultados. Abreviaturas= ACC: Adscripción crono-cultural; E: Excavación; EC: Estructura de combustión; ES: Esqueletos silíceos por PS; H: Período Histórico; IDS: Índice de Diversidad de Simpson; MB: Muestras blancas; PAT: Período Alfarero Temprano; PT: Período Alfarero Tardío; PE: Perfil estratigráfico; UE: Unidad Estratigráfica; PO: Peso original; PL: Peso en lámina; T: Tafonomización.
| Muestra | Unidad | UE | ACC | Profundidad | Muestreo | Origen | PO (g) | PL (mg) | N/fito x g/AIF | ES | T % | IDS |
| P1.1 | P1 | 1-2 | H | 0-10 cm | PE | MB | 4,006 | 0,50 | 24.757 | 8 | 29 | 0,88 |
| P1.2 | P1 | 3 | H | 10-20 cm | PE | MB | 4,004 | 0,80 | 28.058 | 6 | 36 | 0,89 |
| P1.3 | P1 | 3 | H | 20-30 cm | PE | MB | 4,004 | 0,80 | 169.827 | 12 | 7 | 0,88 |
| P1.4 | P1 | 3 | H | 30-40 cm | PE | MB | 4,005 | 0,70 | 118.712 | 22 | 2 | 0,86 |
| P1.5 | P1 | 4 | H | 40-50 cm | PE | MB | 4,003 | 0,90 | 98.852 | 33 | 6 | 0,85 |
| P1.6 | P1 | 4 | H | 50-60 cm | PE | MB | 4,131 | 0,80 | 86.502 | 28 | 8 | 0,86 |
| P1.7 | P1 | 4 | H | 60-70 cm | PE | MB | 4,004 | 1,70 | 219.527 | 8 | 11 | 0,88 |
| P1.8 | P1 | 5 | PT | 70-80 cm | PE | MB | 4,046 | 0,80 | 728.373 | 2 | 7 | 0,8 |
| P1.9 | P1 | 5 | PT | 80-90 cm | PE | MB | 4,026 | 0,70 | 63.914 | 29 | 11 | 0,87 |
| P1.10 | P1 | 5 | PT | 90-100 cm | PE | MB | 4,004 | 1,60 | 131.436 | 35 | 5 | 0,84 |
| P1.11 | P1 | 5 | PT | 100-110 cm | PE | MB | 4,033 | 0,90 | 665.498 | 42 | 5 | 0,83 |
| P1.12 | P1 | 5 | PT | 110-120 cm | PE | MB | 4,008 | 0,50 | 307.737 | 41 | 10 | 0,85 |
| P1.13 | P1 | 5 | PT | 120-130 cm | PE | MB | 4,002 | 1,00 | 90.498 | 33 | 21 | 0,9 |
| P1.14 | P1 | 5 | PT | 130-140 cm | PE | MB | 4,019 | 1,30 | 102.844 | 23 | 26 | 0,87 |
| P1.15 | P1 | 5 | PT | 140-150 cm | PE | MB | 4,038 | 1,00 | 1.049.669 | 67 | 5 | 0,84 |
| P1.16 | P1 | 9.2 | PAT | 150-160 cm | PE | MB | 4,008 | 1,40 | 559.488 | 45 | 28 | 0,84 |
| P1.17 | P1 | 10 | PAT | 160-170 cm | PE | MB | 4,039 | 1,50 | 208.696 | 24 | 19 | 0,85 |
| P1.18 | P1 | 10 | PAT | 170-180 cm | PE | MB | 4,012 | 0,80 | 397.284 | 30 | 25 | 0,83 |
| P1.19 | P1 | 10 | PAT | 180-190 cm | PE | MB | 4,009 | 0,70 | 178.345 | 32 | 26 | 0,81 |
| P1.20 | P1 | 10 | PAT | 190-200 cm | PE | MB | 4,045 | 0,80 | 229.226 | 70 | 23 | 0,83 |
| P1.15b | P1 | 9.1 | PAT | 140-150 cm | PE | EC grande, ceniza | 4,099 | 0,70 | 262.021 | 69 | 57 | 0,9 |
| P2.6 | P2 | 5 | PT | 50-60 cm | E | Lente ceniza | 0,731 | 1,10 | 2.036.314 | 4 | 9 | 0,86 |
| P2.10 | P2 | 7 | PT | 90-100 cm | E | EC, ceniza | 4,047 | 0,80 | 1.221.587 | 24 | 31 | 0,87 |
| P2.11 | P2 | 8.1 | PT | 100-110 cm | E | EC grande, ceniza | 4,017 | 0,60 | 51.093 | 73 | 51 | 0,88 |
| Muestra | Unidad | UE | ACC | Profundidad | Muestreo | Origen | PO (g) | PL (mg) | N/fito x g/AIF | ES | T % | IDS |
| P2.12R2 | P2 | 8.2 | PT | 110-120 cm | E | EC en cubeta, ceniza | 4,019 | 0,70 | 160.992 | 34 | 43 | 0,85 |
| P2.13b | P2 | 8.3 | PAT | 120-130 cm | E | EC grande, ceniza | 1,600 | 0,50 | 1.547.738 | 36 | 41 | 0,86 |
| P2.9R1 | P2 | 6-7 | PT | 80-90 cm | E | EC, termo-alterado | 4,005 | 0,60 | 167.377 | 34 | 27 | 0,81 |
| P3.5 | P3 | - | - | 40-50 cm | E | EC, termo-alterado | 4,042 | 0,50 | 9.018.750 | 15 | 22 | 0,84 |
| P3.5b | P3 | - | - | 40-50 cm | E | EC, termo-alterado | 4,043 | 0,80 | 3.051.623 | 12 | 21 | 0,79 |
| P2.13(1) | P2 | 8.2 | PT | 120-130 cm | E | MB, asoc. semillas carbonizadas | 4,023 | 0,60 | 271.561 | 59 | 35 | 0,85 |
| P2.14(2) | P2 | 8.3 | PAT | 130-140 cm | E | MB, asoc. tubos de pipa | 4,014 | 0,80 | 209.798 | 61 | 26 | 0,8 |
Cuantificación
La abundancia se presenta de acuerdo a la concentración de silicofitolitos por gramo de AIF (Albert & Weiner, 2001). Este método permite cuantificar la cantidad de fitolitos con respecto a la fracción mineralógica de la muestra, que suele ser más estable en comparación a la proporción de materia orgánica u otros componentes del sedimento.
Análisis estadístico
Para comparar las muestras, aplicamos en primer lugar estadísticos descriptivos (frecuencia absoluta y relativa), Índice de Diversidad de Simpson (Marston, 2014), considerando variables nominales, sobre el Grand-Total de fitolitos. No obstante, para la comparación entre dicotiledóneas y monocotiledóneas, solo se tomaron en cuenta categorías relevantes a nivel taxonómico (Tabla 2). Adicionalmente, desarrollamos el Análisis de Correspondencia (AC), de manera de hacer visibles posibles patrones de la muestra en estudio, pues este tipo de análisis multivariado simplifica la visualización de los datos en dos dimensiones (Barceló, 2007; Smith, 2014; Baxter, 2015). Antes de llevar a cabo el AC, se desarrolló el test de Chi-cuadrado en base a la hipótesis nula de que los morfotipos están distribuidos igualmente en las muestras (Shennan, 1992; Barceló, 2007). Con el fin de utilizar solo categorías estadísticamente representativas, se agruparon morfotipos en categorías más amplias, aunque sin perder su valor anato-taxonómico y excluyendo las categorías con un conteo menor al 1% de la muestra. Todos los análisis y figuras se desarrollaron en Microsoft Excel y Past3.
Resultados
Todas las muestras presentaron silicofitolitos y tejidos silicificados, además de diferentes restos de plantas como tejidos quemados y microcarbones; y otros organismos de sílice biogénico como diatomeas y espículas de espongiarios. Todos estos se encontraban relativamente en buen estado de conservación.
Un 87% del total de silicofitolitos fueron determinados (n= 6864), clasificándolos en 44 morfotipos, y cuatro categorías más amplias con valor taxonómico que no pudieron clasificarse en un morfotipo particular, ya sea por su posición en el portaobjeto o por su estado de tafonomización: 1) Monocotiledóneas no-determinadas, 2) células cortas no-determinadas, 3) Dicotiledóneas no-determinadas y 4) células epidérmicas Dicotiledóneas no-determinadas. Por otra parte, el 13% del total de silicofitolitos que no pudo ser agregado a categorías morfo-tipológicas concretas, se considera "no-determinado" (n= 1051).
Diversidad
La diversidad fue calculada con el Índice de Simpson (Tabla 1), bajo la hipótesis de que las muestras de estructuras de combustión serían más diversas. Se observa una alta diversidad en todas las muestras, no obstante, las muestras de rasgos, en específico de ceniza (0,85-0,9), se presentan ligeramente más diversas que las muestras blancas (0,79-0,85).
Tabla 2. Clasificación de silicofitolitos de la Cueva de los Catalanes. Abreviaturas= CL: célula larga; CB: célula buliforme; CC: célula corta; G: globular; CE: célula epidérmica; ND: silocofitolito no-determinado; PS: phytolith sum.
Abundancia y preservación
De acuerdo a la abundancia de silicofitolitos (Tabla 1, número de fitolitos por gramo de AIF), los resultados siguen cierto patrón. Al comparar los valores mínimos y máximos, y la mediana entre muestras de rasgos y muestras blancas (Tabla 3), se observa que los rasgos presentan los picos más altos de todo el conjunto, correspondiendo estos, en primer lugar, a los rasgos de sedimento termo-alterado de P3, seguidos de las muestras de ceniza de distintos estratos de P2 (P2.6, P2.10, P2.13b). Cabe destacar que, aunque muy por debajo de los máximos presentados por las muestras de rasgos, las muestras blancas mostraron sus mayores picos en los niveles prehispánicos.
En relación a la abundancia de tejidos o esqueletos silíceos (Tabla 1), se observa una tendencia de aumento hacia los niveles más profundos (prehispánicos). Al comparar los valores mínimos, máximos y la mediana, se observa que los valores son bastante similares entre muestras blancas y muestras de rasgos (Tabla 3). En el caso de las muestras de rasgo, las mayores abundancias están representadas por muestras de ceniza (P2.11, P1.15b) de las capas 8 y 9 (período Alfarero temprano); mientras en las muestras blancas las mayores abundancias también se encuentran en estas capas (P1.15, P2.13, P2.14, P1.16), además de una muestra de la capa 10 (P1.20). Cabe destacar que este último estrato se asienta en la roca madre de la cueva, por lo cual sospechamos que esta abundancia se debe a migración vertical.
Finalmente, la tasa de tafonomización de los silicofitolitos resultó ser relativamente baja (Fig. 3, Tabla 1), y los mayores picos se presentan en las muestras de rasgos de ceniza (P1.15b, P2.11, P2.12R2, P2.13b). Entre las muestras blancas, la mayor parte presenta menor tafonomización, aunque ésta tiende a ser más alta en los niveles superiores (histórico-subactuales; P1.1, P1.2), por una parte y, por otra, entre las capas 5, 8, 9 y 10 (niveles prehispánicos; P1.13, P1.14, P2.13, P2.14, P1.16, P1.17, P1.18, P1.19, P1.20). En otras palabras, en los niveles más profundos tiende a haber una mayor tafonomización.
Espectro
Los morfotipos de este análisis han sido propuestos desde trabajos que han investigado la flora de distintas partes del mundo (Fig. 4). La mayor parte de éstos corresponden a Angiospermas, las que se dividen en dos clases según el esquema de clasificación de Cronquist (1981): monocotiledóneas (Liliopsida) y dicotiledóneas (Magnoliopsida), el cual se ha reorganizado en los últimos años (ej. APG, 2016). En el presente estudio seguimos utilizando la clasificación clásica por su valor en el análisis de silicofitolitos y, en especial, en el estudio de estructuras de combustión, asumiendo que dentro de las dicotiledóneas se encuentran las plantas leñosas (combustible más común).
Con todo, se presentaron 24 morfotipos de monocotiledóneas y 21 de dicotiledóneas (Tabla 2). Entre las monocotiledóneas, los morfotipos más abundantes corresponden a células cortas (Poaceae), células largas, y células buliformes. Entre las primeras, destacan los morfotipos trapezoidal, rondel y bilobado; mientras que entre las células largas destacan las de borde psilado, sinuoso, almenado, columnado (estas tres las simplificamos en un solo morfotipo sin-alm-col), equinado y células dendríticas (estas últimas diagnósticas de Poaceae). El morfotipo más abundante de las células buliformes es el cuneiforme. Los estomas y las papilas se presentaron en menor frecuencia. Conviene destacar en esta categoría los conos de Cyperaceae que, aunque presentaron una relevancia menor en términos de frecuencia, presentan valor diagnóstico taxonómico a nivel de familia.
Tabla 3. Abundancia de fitolitos y tejidos con los valores mínimos y máximos, y la mediana.
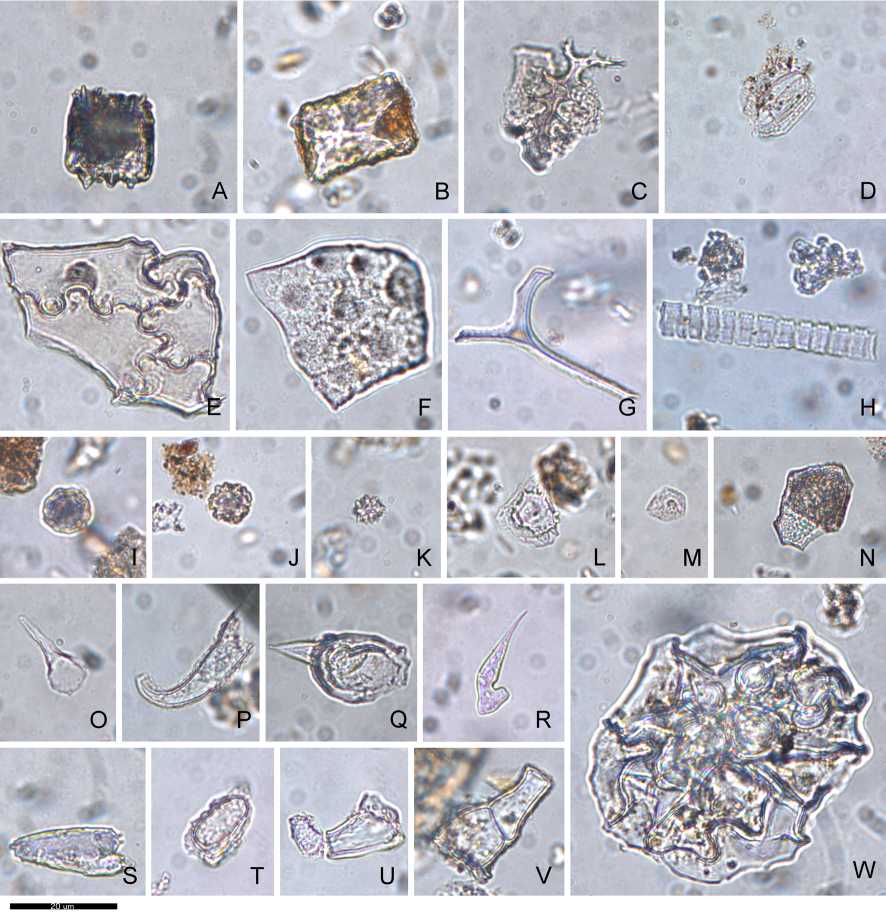
Fig. 3. Silicofitolitos con daño. A: Daño químico (muestra P1.13). B: Daño mecánico (muestra P1.11). Escala= 20 pm.
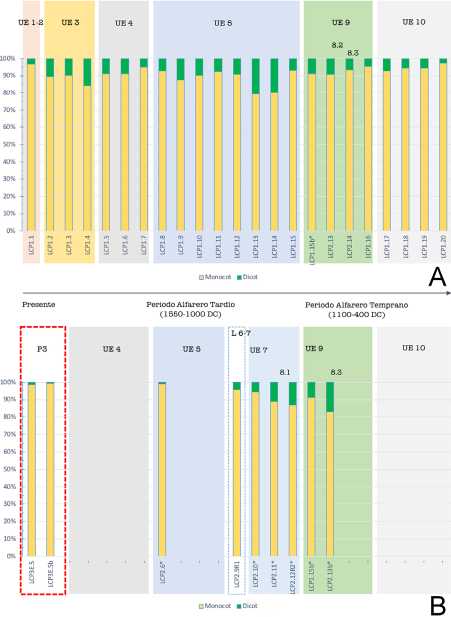
Fig. 4. Morfotipos en sedimentos de la Cueva de los Catalanes. A: Célula buliforme paralelepípeda. B: Blocky. C: Dendrítico. D: Estoma. E: Célula epidérmica "jisaw puzzle". F: Célula epidérmica. G: Esclereida. H: Traqueida. I: Globular granulado. J: Globular verrugoso. K: Globular equinado. L: Papila. M: Cono Cyperaceae. N: Célula epidérmica poliédrica escrobiculada. O: Pelo acicular (ND). P: Pelo acicular unciforme (ND). Q: Tricoma 1. R: Tricoma 3. S: Tricoma 4. T: Base de pelo ND. U: Célula de pelo 8. V: Célula de pelo 10. W: Base de pelo multicelular (dicotiledónea). Escala= 20 pm.
En relación a las dicotiledóneas, los morfotipos más abundantes son el globular granulado y las células epidérmicas. Entre estas últimas se destacan los morfotipos poligonal y jigsaw. Cabe destacar que las esclereidas fueron contabilizadas dentro de esta categoría porque se presentan mayormente en esta clase de plantas, en ciertos climas son incluso diagnósticas de taxa arbóreos (Piperno, 1988).
Adicionalmente, se presentó un morfotipo de probable gimnosperma ("rectangular pitted"; Carnelli et al., 2004), dos morfotipos correspondientes a elementos vasos (traqueidas y células vasculares), además de cuatro morfotipos de células de pelo aparentemente sin valor taxonómico.
Dicotiledóneas versus monocotiledóneas: A continuación, interesó evaluar la proporción de plantas dicotiledóneas en relación a monocotiledóneas por muestra; de acuerdo a la hipótesis de que las muestras de rasgos presentarían una mayor proporción de dicotiledóneas que las muestras blancas. Sin embargo, como se observa (Fig. 5A-B), todas las muestras tienen una ostensible mayor proporción de morfotipos de monocotiledóneas (esto tiene una explicación que será abordada en la discusión). No obstante, mientras los rasgos de niveles menos profundos suelen presentar una muy alta proporción de morfotipos de monocotiledóneas, las muestras de rasgos de las capas 8 y 9 presentan las mayores proporciones de dicotiledóneas, correspondiendo todas a rasgos de ceniza (P2.11, P2.12R2, P1.15b, P2.13b). En cuanto a las muestras de P3 (sedimentos termo-alterados), éstas presentan una evidente mayor proporción de monocotiledóneas, incluso al compararlas con las muestras sedimentarias, algo similar sucede con P2.6 (rasgo de ceniza).
Partes anatómicas: Aquí, interesó evaluar los órganos de plantas representados en el registro, en función de lo cual se excluyeron algunos morfotipos que no entregaban información relevante a este respecto. Para este ejercicio se consideraron solo 31 morfotipos, correspondientes a un 40,63% del grand-total (n= 3216) (Fig. 6A). Los morfotipos de monocotiledóneas presentes en las muestras corresponden a fitolitos de tallo-hoja (células largas, tricomas), hoja (células buliformes y estomas) e inflorescencia-fruto (dendríticos y papilas). La categoría tallo-hoja presentó la distribución más importante en el conjunto de muestras, seguida de la categoría hojas. Cabe destacar que estos morfotipos resultaron tener mayor valor diagnóstico de distintas partes anatómicas, a diferencia de los morfotipos de dicotiledóneas, los que no diferencian categorías anatómicas tan precisas.
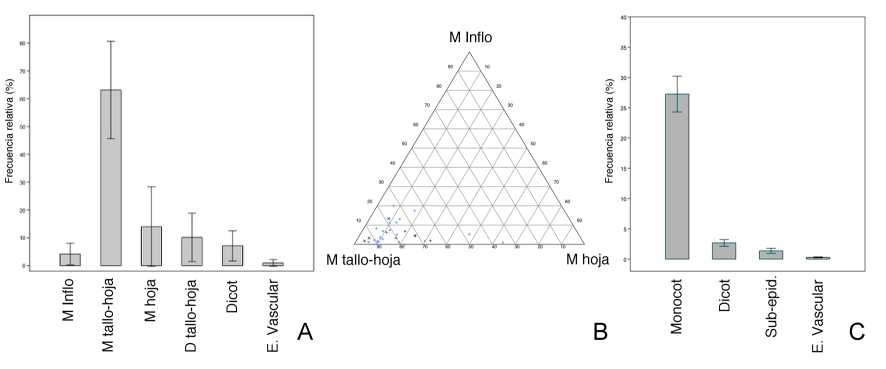
Fig. 5. Proporción de monocotiledóneas versus dicotiledóneas. A: Proporción en las muestras sedimentarias. B: Proporción en las muestras de estructuras (símbolo *: muestras de ceniza). Abreviaturas= Monocot: Monocotiledóneas; Dicot: Dicotiledóneas.
Los morfotipos de plantas dicotiledóneas, por su parte, se clasificaron en fitolitos de hoja-tallo (en herbáceas) o solo de hoja (en leñosas), dicotiledónea de proveniencia anatómica indeterminada (morfotipos publicados de taxa leñosos y/o arbóreos, ej. globular granulado; Bremond et al., 2005), y elementos de vasos. Si bien estos últimos no solo se presentan en plantas dicotiledóneas, se registran de manera más común en estas, así como en abundancia en sedimentos de bosques templados de ciertas regiones, por lo que tentativamente los agrupamos dentro de esta clase (Tsartsidou et al., 2007; Roa, 2016; Gao et al, 2018). Los morfotipos de tallo-hoja y hoja fueron los más abundantes, seguidos por la categoría de dicotiledónea.
Los morfotipos de dicotiledóneas sólo entregaron información de las hojas (y tallos) principalmente, por lo cual se decidió comparar las muestras en base solo a la información entregada por los morfotipos de monocotiledóneas en un diagrama ternario (Fig. 6B). La hipótesis que seguimos aquí es que sería más probable encontrar morfotipos de frutos e inflorescencias de gramíneas en los períodos más tardíos e incluso post-contacto; esto bajo el supuesto de que durante el período Alfarero tardío se habría intensificado el uso de granos y ya en momentos post-contacto ingresarían cereales europeos. En el diagrama se observa que la mayor parte de las muestras se agrupa hacia el vértice de tallo-hoja, y dos muestras se separan del resto presentando una mayor proporción de morfotipos de hoja (P1.1 y P1.2), correspondientes a niveles subactuales. A su vez, destacan tres muestras por alejarse del vértice hacia los morfotipos de fruto e inflorescencias (P1.6, P1.7, P1.10), los que se corresponden con niveles de los períodos Alfarero tardío e histórico.
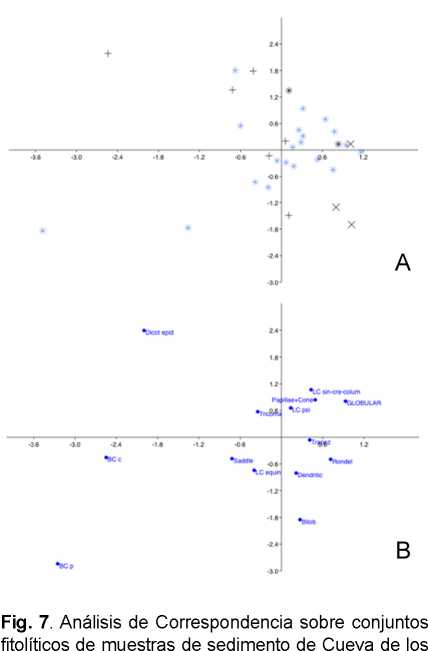
Fig. 6. Proporciones de morfotipos de fitolitos y esqueletos silíceos. A: Proporciones de las clases de morfotipos de los fitolitos de relevancia anatómica y taxonómica. B: Diagrama ternario comparativo entre distintas partes de plantas de monocotiledóneas (símbolos= *: muestras blancas; +: cenizas; x: sedimentos termo-alterados). C: Proporciones de las clases de morfotipos de los esqueletos silíceos de relevancia anatómica y taxonómica. Abreviaturas= M: Monocotiledónea; Inflo: inflorescencia; D: Dicotiledónea; E. Vascular: elemento vascular; Monocot: Monocotiledónea; Dicot: Dicotiledónea; Sub-epid.: sub-epidérmico.
Tejidos silicificados: En relación a los esqueletos silíceos (Fig. 6C), se registró tejido epidérmico, tejido subepidérmico y elementos vasos. De ellos, el tejido epidérmico es taxonómicamente relevante, pudiendo diferenciar entre las clases monocotiledónea y dicotiledónea (Ortega et al, 1993), correspondiendo en su gran mayoría a las primeras. En cuanto a las estructuras subepidérmicas, éstas no permiten su asignación a las clases taxonómicas, no obstante es posible que en su mayoría correspondan a plantas monocotiledóneas, dada su significativa tasa de silicificación.
Análisis estadístico multivarado
Como se explicó con anterioridad, el análisis se desarrolló sobre un total de 14 morfotipos (n= 5631 fitolitos).
El análisis de Chi-cuadrado, cuyo valor p (4,1542 x 10-192), permitió rechazar la hipótesis nula, indicando que las variables morfotipos y muestras están asociadas, aunque esta asociación es precaria (V de Cramer= 0,16).
Los resultados del Análisis de Correspondencia muestran que el conjunto se ordenó en13 dimensiones, el cual es un número elevado. De todos modos, las dos primeras dimensiones alcanzaron una inercia acumulada de 51,164%, correspondiente al nivel de explicación del conjunto, lo cual se considera aceptable.
Ahora bien, al interpretar el biplot (Fig. 7A) observamos en primer lugar que, a lo largo del eje x (dimensión 1) la mayor parte de las muestras se ubica en los cuadrantes derechos, y al agregar el eje y (dimensión 2) la mayoría se concentra en el cuadrante superior derecho; esto es válido para la mayor parte de las muestras blancas. En segundo lugar, se observa que las muestras de ceniza (P1.15b, P2.10, P2.11, P2.13b) tienden a segregarse hacia el lado izquierdo (eje x) y en solo dos casos (P2.12R2, P2.6) se mantienen en el lado derecho cercanos a 0 en el eje x. El grupo mayoritario del conjunto se separa en dirección superior izquierda al agregar la dimensión 2, en la dirección del morfotipo de células epidérmicas de dicotiledónea. En tercer lugar, en cuanto a las muestras de rasgos de sedimento termo-alterado, se observa que en el eje x se ubican hacia la derecha, pero al agregar la dimensión 2 las muestras de los rasgos de P3 (P3E.5 y P3E.5b) se segregan hacia el cuadrante izquierdo e inferior, donde se asocian a una muestra de rasgo de ceniza (P2.6); resulta interesante que este conjunto se mueve en dirección opuesta al de la mayor parte de los rasgos de ceniza descritos anteriormente. Finalmente, otra distribución llamativa corresponde a aquélla de los niveles superiores de P1 (P1.1, P1.2, P1.3), las cuales se escapan en distintos grados hacia el cuadrante inferior izquierdo, en dirección de los morfotipos de células buliformes (Fig. 7B).
En el resto de los casos, la asociación entre muestras y morfotipos no es clara lo cual puede deberse a que se pierde una buena parte de la información al considerar solo dos dimensiones (de un total de 13).
Catalanes. A: Biplot Alas de las muestras blancas y rasgos. B: Biplot columnas de morfotipos (símbolos= *: muestras blancas; +: cenizas; x: sedimentos termo-alterados). Abreviaturas= Dicot epid: célula epidérmica de dicotiledónea; LC sin-cre-collum: células largas sinuosa-almenada-columnada; LC psi= células largas psiladas; LC echin= célula larga equinada; Trapez: célula corta trapezoidal; Bilob: célula corta bilobada; BC c: célula buliforme cuneiforme; BC p: célula buliforme paralelepípeda.
Discusión
Las muestras de Cueva de los Catalanes contienen una abundancia significativa de fitolitos, como se esperaría para un registro vegetal de origen antropogénico (Hansen, 2001; Madella & Lancelotti, 2012). En relación a las muestras blancas en la gradiente estratigráfica (P1), las abundancias de silicofitolitos son más significativas a partir de una profundidad de 70 cm (capas 8, 9, 10), sugiriendo que los niveles prehispánicos tuvieron un input vegetal mayor, lo que puede deberse al carácter de la ocupación (más intensa, más prolongada, o con mayor procesamiento de plantas). Los tejidos también presentan relativamente mayores abundancias en los niveles prehispánicos (capas 5, 8, 9, 10). La ocurrencia de tejidos o agregados silíceos pueden dar cuenta de la estabilidad del depósito y de la degradación de material vegetal in situ (Zurro, 2006; Shillito, 2011; Shillito & Matthews, 2013). En este sentido, las mayores abundancias de tejidos en estas capas de cronología prehispánica, darían cuenta del procesamiento y descarte de plantas en el sitio.
Por el contrario, al tomar en cuenta las muestras más superficiales en la gradiente estratigráfica (P1.1, P1.2, P1.3), éstas se comportan diferente en relación a algunos parámetros; mostrando una baja abundancia de fitolitos aislados y tejidos, una mayor tafonomización, además de una composición del conjunto fitolítico distinta: presentaron las mayores proporciones de fitolitos hojas de monocotiledóneas (células buliformes y estomas). Estas muestras corresponden a niveles de estabulación recientes de la cueva (siglos XX y XXI), de tal manera que una mayor cantidad de hojas en este contexto podría deberse a heno y/o a las fecas de herbívoros (Shillito & Matthews, 2013). Por lo cual, vemos que el registro fitolítico de estas actividades es distinguible del resto de las ocupaciones.
Volviendo al objetivo central del trabajo, se pudo determinar que las muestras de rasgos son más heterogéneas que las muestras blancas y que, en general, presentan mayores picos de abundancia de silicofitolitos aislados. Asimismo, en el conjunto de rasgos se observó que las muestras más abundantes en silicofitolitos, presentaron menos agregados silíceos, y viceversa. En este sentido, se puede proponer que las áreas con un input vegetal significativo estarían presentando una estabilidad menor, ya sea pre-depositacional (un procesamiento de plantas más intensivo) o post-depositacional (mayor remoción de los depósitos a causa de actividades antrópicas más intensivas).
Si nos detenemos en el contexto de estudio, desde la etnohistoria y la etnografía sabemos que en la sociedad mapuche los fogones no solo fueron utilizados para comer, sino también como puntos de encuentro donde la gente podía pasar la noche trabajando y compartiendo con la familia, así como atestigua un relato de Lanco: "[las mujeres] en la noche, nos amanecíamos hilando" (Egert & Godoy, 2008).
Por otra parte, la tafonomización de los fitolitos en las muestras de rasgos de ceniza es relativamente mayor a las muestras sedimentarias; lo anterior es claro en la muestra P 1.15b (ceniza), cuya mayor tafonomización contrasta al ser comparada con el resto de las muestras blancas de P1. Incluso, la tafonomización tiende a incrementarse hacia los niveles más profundos. Este mayor daño tafonómico podría relacionarse a la composición de las cenizas con abundante carbonato de calcio, el que representa un medioambiente menos proclive para la preservación de ópalos de sílice (Weiner, 2010).
Dentro de las muestras de rasgos, llama la atención el conjunto fitolitológico de las muestras de sedimentos termo-alterados de P3. Éstas resaltan por su mucho mayor abundancia de fitolitos y mucho menor presencia de tejidos, una notoria mayoría de morfotipos de plantas monocotiledóneas, además de presentar una menor tafonomización. Asimismo, estas muestras de P3 se comportaron de manera similar junto con una muestra de rasgo de ceniza (P2.6), lo que podría estar relacionado a una segregación cronológica, dado que estos rasgos se encuentran en niveles menos profundos y, por consiguiente, menos antiguos que el resto de los rasgos.
Dentro del conjunto de estructuras de combustión hay distintas configuraciones (Tabla 1): rasgos de ceniza extendidos (P1.15b, P2.11, P2.13b), de tamaño medio (P2.10), en cubeta (P2.12R2, llamado "cenicero" en publicaciones previas; Menghin, 1959-1960; Berdichewsky, 1968) y un pequeño lente (P2.6). En relación al conjunto fitolitológico no se observaron diferencias entre los rasgos atribuibles a su morfología. Lo anterior se correlaciona con observaciones etnográficas donde las características morfológicas de los fogones no se relacionan a función o usos particulares, tampoco a la duración de la ocupación (Galanidou, 2000). En este sentido, las distintas morfologías de las estructuras de ceniza pueden no estar relacionadas a conjuntos de fitolitos específicos, ya que no existiría una necesaria correlación entre un uso particular y la morfología.
Así, a pesar de que no encontramos regularidades del registro en relación a la forma, podemos expresar que el espectro de fitolitos no nos permitió definir usos específicos de cada uno de estos rasgos, ya que éstos se derivarán de la interpretación del yacimiento y sus asociaciones contextuales. Sin embargo, podemos corroborar que estos espacios concentraron actividades relacionadas al uso de plantas, que permitieron mostrar un input vegetal significativo a la vez que heterogéneo. En efecto, se pudo atestiguar la presencia de distintos órganos de distintas clases de plantas, lo que permitirá enunciar hipótesis guía de otros trabajos, donde se podrán mejorar las técnicas de recuperación de las muestras acordes a una pregunta específica de investigación.
En referencia a la mayor proporción de plantas monocotiledóneas versus dicotiledóneas en el conjunto de muestras, incluso en aquellas muestras de rasgos, esto se explica en parte porque la acumulación de sílice está genéticamente determinada en las plantas. En este sentido, las monocotiledóneas (especialmente las familias Poaceae y Cyperaceae) suelen presentar una cantidad considerable de taxones acumuladores de Si y, por el contrario, muchas dicotiledóneas corresponden a taxones no-acumuladores de Si (Ma, 2003). Adicionalmente, se ha propuesto que cuando los taxones acumulan sílice, las monocotiledóneas lo hacen entre un 20-40% más que otras plantas (Albert & Weiner, 2001). A pesar de que esta subrepresentación de las plantas dicotiledóneas en el registro fitolitológico se considera un obstáculo al comparar ambas clases de plantas, es interesante observar que el hecho de que los rasgos de ceniza presenten evidencias de plantas monocotiledóneas, significa que este tipo de plantas está siendo quemado en la estructura de combustión, ya sea como iniciadora, como combustible, o como un residuo de actividades en torno al hogar.
La recolección de leña en Araucanía fue una actividad que se podía realizar sin dificultad en los alrededores de los asentamientos, tal como atestiguaron los cronistas (Núñez de Pineda y Bascuñán, 1863 [1673]; Góngora de Marmolejo, 1960[1575]), sin embargo, estos resultados llevan a pensar que la disponibilidad de leña no necesariamente dirigió un uso solo de plantas leñosas como combustible primario, lo cual ha sido sugerido en otros trabajos (Aldeias, 2017). Esto se hace más claro al observar que la mayor parte de las evidencias corresponderían a tallos y hojas de monocotiledóneas, es decir, que la planta completa fue acarreada y quemada en la estructura.
Un comentario acerca de las diferencias en la secuencia de ocupación prehispánica
Finalmente, nos referiremos a ciertas diferencias entre los conjuntos fitolitológicos de los niveles relativos al período Alfarero temprano (PAT) y aquellos del Alfarero tardío (PT). Esto a través de las muestras blancas recuperadas de un perfil estratigráfico, considerando las muestras del PAT (P1.16, P1.17, P1.18, P1.19, P1.20) y del PT (P1.8, P1.9, P1.10, P1.11, P1.12). Como veremos, a pesar de que las muestras blancas corresponden a áreas donde no fue posible distinguir a primera vista el uso de plantas (como sí es el caso de las muestras de rasgos), debe considerarse el supuesto de que los sedimentos de los sitios arqueológicos contienen información valiosa acerca del procesamiento y los restos de las plantas (si fueron utilizadas), las que son invisibles al ojo desnudo (Juan i Tresserras, 1997).
Durante el PAT, la menor heterogeneidad de las muestras sugiere una mayor regularidad en la ocupación o en la estructuración del espacio. Una significativa agregación de tejidos (mayores abundancias de esqueletos de sílice), demuestra además una mayor estabilidad de los depósitos, lo que sugiere una mayor descomposición de las plantas in situ, o bien un menor transporte o remoción de los sedimentos. Lo anterior es coherente con posibles ocupaciones más breves, aunque redundantes, y posiblemente ligadas a un patrón estacional del uso del espacio.
En relación al espectro, una mayor proporción de plantas monocotiledóneas se sustenta en una mayor proporción de estos morfotipos (especialmente célula larga psilada, ubicada en distintas familias y partes anatómicas de esta clase de plantas).
Lo cual sugiere el uso de plantas herbáceas provenientes de praderas o humedales, ya que prácticamente las únicas monocotiledóneas del bosque son las quilas o colihues (Chusquea spp.). Por su parte, la ausencia de células dendríticas en estas muestras es llamativa, dando cuenta de una virtual ausencia de inflorescencias-cascarinas de cereales silvestres en el registro. Combinando los datos, se podría sugerir que los tallos y hojas de gramíneas están siendo mayormente utilizados que las inflorescencias-granos, posiblemente a causa de una selección de tallos/hojas para actividades específicas, o bien por el uso estacional de la cueva durante el período otoño-invierno. No obstante, aunque los dendríticos no aparecen en las muestras blancas, sí se encuentran en el registro de algunas muestras de rasgos de ceniza de posible cronología temprana (P 1.15b y P2.13b), lo cual nos permite aducir el uso de inflorescencias solo en el contexto de fogones durante el PAT. Adicionalmente, al examinar la distribución de otros morfotipos relativos a la inflorescencia (como papilas), se observa que están en las muestras blancas de ambos períodos. Es posible entonces que el uso/procesamiento de cereales silvestres (como Bromus spp.) fue menor en este período.
Durante el PT se observan claras diferencias en las abundancias entre las muestras (mayor heterogeneidad), la que pudiera ser una consecuencia del carácter de la ocupación. En este sentido, en base al registro fitolitológico, sugerimos que la ocupación pudo ser menos intensiva o menos redundante en el tiempo. En segundo lugar, la proporción más significativa de plantas dicotiledóneas en el PT (que se hace más evidente al observar el aumento de los morfotipos globulares, en especial el globular granulado que da cuenta de especies leñosas o arbóreas, distribuido en distintas partes anatómicas), sugiere que estas plantas tuvieron un uso más intensivo en este período. Desde la etnohistoria se ha podido definir un amplio uso del espectro de plantas disponible por parte de las poblaciones reche-mapuche, incluyendo la talla en madera, la herbolaria, o el procesamiento de cultivos, el que pudiera tener su antecedente durante este período. Lo anterior, unido a la menor agregación de los restos silíceos (menor cantidad de esqueletos de sílice), puede interpretarse como una menor estabilidad de los depósitos (mayor remoción y transporte de sedimentos) y/o procesamiento de los restos de plantas (ej. molienda). De todas maneras, otra hipótesis se deriva de la producción de sílice en las plantas, ya que una menor proporción de esqueletos podría relacionarse a una menor presencia de plantas acumuladoras de sílice (coincidente con la relativa menor proporción de monocotiledóneas), o a condiciones de menor humedad en la vida de la planta.
Durante este período destaca además la presencia de las células dendríticas3 en las muestras blancas, a diferencia del PAT. Como dijimos anteriormente, dicha evidencia sugiere el uso de los granos de cereales silvestres como aquellos del género Bromus; los que pueden ser recolectados durante la primavera. El uso de los granos de estas plantas como "famine food" pudo tener una mayor relevancia durante el PT, cuando estos grupos estaban más adaptados a un modo de vida agricultor, con un mayor consumo de granos de maíz o quínoa, lo que pudo incentivar una mayor selección de granos de cereales silvestres cuando las reservas de grano cultivado se agotaban. En este sentido, en la cosmovisión mapuche contemporánea pewü o primavera se concibe como un tiempo de escasez, presentándose justo antes de las cosechas del verano. Por otra parte, existen datos del uso de granos de Bromus mango E. Desv. en el sur de Chile hasta los siglos XIX y XX (Gay, 1865; Coña, 2010[1930]; Hilger, 2015). Al respecto, destacamos que en las muestras del PH que siguen a los niveles del PT (P1.7, P1.6, P1.5), se observa un aumento definitivo de los dendríticos, disminuyendo considerablemente las formas globulares; en el diagrama ternario las muestras P1.7, P1.6 y P1.10 se diferenciaron del resto por la presencia de dendríticos. Lo anterior podría interpretarse como un cambio dado por la adopción de cereales cultivados a partir del contacto con europeos (presentes en el registro carpológico). Si consideramos que las muestras de rasgos más superficiales que se comportan de una manera similar (P3 y P2.6), provendrían de niveles post-contacto, vemos clara la intensificación en el uso de plantas monocotiledóneas, presentándose además un input vegetal mucho mayor.
En conclusión, pensamos que el registro fitolitológico del PT, estaría mostrando una diversificación e intensificación en el uso de plantas en la cueva, aunque de manera más heterogénea en el tiempo y espacio.
Conclusiones
En referencia a los objetivos del trabajo, los resultados permiten sostener la relevancia del análisis de fitolitos como una herramienta útil y complementaria para entender el manejo del fuego, además del consumo de plantas, en la región de estudio. Este tipo de análisis presenta un escaso desarrollo, aunque un amplio potencial aún inexplorado.
En segundo lugar, desde las características del conjunto fitolítico fue posible observar diferencias en las muestras de ceniza en comparación al resto del conjunto: en efecto, se comprobó una mayor abundancia y diversidad de morfotipos en estas muestras. En cuanto al espectro, es llamativa la importante presencia de monocotiledóneas en este tipo de rasgos.
En tercer lugar, se propone una segregación entre los rasgos más tardíos (posiblemente post-contacto) y los más tempranos (prehispánicos), aunque no se observaron diferencias claras entre las ocupaciones PAT y PT. No obstante, al considerar el conjunto global de muestras (incluidas las muestras blancas), destacaron señales específicas para las distintas ocupaciones. Destaca, por ejemplo, el mayor input vegetal en las ocupaciones prehispánicas.
En cuarto lugar, para futuros trabajos se deberá enfatizar en una toma de muestras con objetivos arqueobotánicos claros. Estas estrategias deberán considerar al menos el contenido de las estructuras de combustión (cenizas), su parte basal (sedimentos termo-alterados), así como muestras blancas en asociación horizontal.
Contribución de los autores
CR y DZ diseñaron la investigación, interpretaron los datos, confeccionaron las figuras y escribieron el manuscrito. CR colectó las muestras y llevó a cabo el análisis.
Agradecimientos
Al Dr. Frances Brnjachs (IPHES-ICREA), Jonas Alcaina (CaSEs, UPF), Josep Vallverdú (IPHES), Laboratorio Biogeopal (IMF-CSIC), Unidad Arqueobotánica (IPHES) y Camila Roa por las imágenes. A las coordinadoras de la Mesa de Comunicaciones Estudios en Patagonia, III Jornadas Argentinas de Etnobiología y Sociedad, La Plata. Este trabajo fue parte de una tesis de maestría financiada por el Proyecto FONDECYT N° 1115039, beca Erasmus+ (International Master in Quaternary and Prehistory, Unión Europea). A los revisores del BSAB.
Bibliografía
ABARZÚA, A. M., A. PICHUNCURA, L. JARPA, A. MARTEL-CEA, M. STERKEN, R. VEGA & M. PINO. 2014. Environmental responses to climatic and cultural changes over the last 26,000 years in Purén-Lumaco valley (38°S). En: DILLEHAY, T. (ed.), The Telescopio Polity. Andean Patriarchy and Materiality, pp. 123-141. Springer, New York. https://doi.org/10.1007/978-3-319-03128-6 ADÁN, L. & R. MERA. 2011. Variabilidad interna en el Alfarero Temprano del centro-sur de Chile: el Complejo Pitrén en el valle central del Cautín y el sector lacustre andino. Chungara 43: 3-23. https://doi.org/10.4067/S0717-73562011000100001 ALBERT, R. M., S. WEINER, O. BAR-YOSEF & L. MEIGNEN. 2000. Phytoliths in the Middle Palaeolithic deposits of Kebara cave, Mt Carmel, Israel: study of the plant materials used for fuel and other purposes. J. Archaeol. Sci. 27: 931-947. https://doi.org/10.1006/jasc.2000.0507 ALBERT, R. M. & S. WEINER. 2001. Study of phytoliths in prehistoric ash layers from Kebara and Tabun caves using a quantitative approach. En: MEUNIER, J. D. & F. COLIN (eds.), Phytoliths: Applications in Earth Sciences and Human History, pp. 251-266. Balkema Publishers, Lisse. https://doi.org/10.1201/9780367800406
ALDEIAS, V. 2017. Experimental approaches to archaeological fire features and their behavioral relevance. Curr. Anthropol. 58 (Supp. 16): S191-S205. https://doi.org/10.1086/691210 ANGIOSPERM PHYLOGENY GROUP. 2016. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 181: 1-20. https://doi.org/10.15468/fzuaam BALL, T., L. VRYDAGHS, I. VAN DEN HAUWE, J. MANWARING & E. DE LANGHE. 2006. Differentiating banana phytoliths: wild and edible Musa acuminata and Musa balbisiana. J. Archaeol. Sci. 33: 1228-1236.
https://doi.org/10.1016/jjas.2005.12.010 BALL, T., R. EHLERS & M. D. STANDING. 2009. Review of typologic and morphometric analysis of phytoliths produced by wheat and barley. Breed. Sci. 59: 505-512. https://doi.org/10.1270/jsbbs.59.505 BARBETTI, M. 1986. Traces of fire in the archaeological record, before one million years ago? J. Hum. Evol. 15: 771-781.
https://doi.org/10.1016/S0047-2484(86)80009-4 BARCELÓ, J. A. 2007. Arqueología y estadística. Introducción al estudio de la variabilidad de las evidencias arqueológicas. Universitat Autónoma de Barcelona, Bellaterra.
BAXTER, M. J. 2015. Exploratory Multivariate Analysis in Archaeology. Percheron Press, New York. https://doi.org/10.2307/j.ctv2sx9gfb BERDICHEWSKY, B. 1968. Excavaciones en la Cueva de los Catalanes (Provincia de Malleco). Boletín de Prehistoria de Chile 1: 33-83.
BERNA, F., A. BEHAR, R. SHAHACH-GROSS, J. BERG, E. BOARETTO, A. GILBOA, I. SHARON, S. SHALEV, S. SHILSTEIN, N. YAHALOM-MACK, J. R. ZORN & S. WEINER. 2007. Sediments exposed to high temperatures: reconstructing pyrotechnogical processes in Late Bronze and Iron Age Strata at Tel Dor (Israel). J. Archaeol. Sci. 34: 358-373. https://doi.org/10.1016/jjas.2006.05.011 BINFORD, L. R., 1991[1983]. La gente en su espacio. En: BINFORD, L. R. (ed.), En Busca del Pasado, pp. 154-203. 2da Ed. Editorial Crítica, Barcelona. BOZARTH, S. 1992. Classification of opal phytoliths formed in selected Dicotyledons native to the Great Plains. En: RAPP, G. & S. MULHOLLAND (eds.), Phytolith Systematics. Emerging Issues, pp. 193214. Springer, Minnesota. https://doi.org/10.1007/978-1-4899-1155-1
BREMOND, L., A. ALEXANDRE, C. HÉLY & J. GUIOT. 2005. A phytolith index of tree cover density in tropical areas: Calibration with Leaf Area Index along a forest-savanna transect in southeastern Cameroon. Glob. Planet. Change 45: 277-293. https://doi.org/10.1016Zj.gloplacha.2004.09.002 CABANES, D., E. ALLUÉ, J. VALLVERDÚ, I. CÁCERES, M. VAQUERO & I. PASTÓ. 2007. Hearth structures and function at level J (50kyr, bp) from Abric Romaní (Capellades, Spain): phytolith, charcoal, bones and stone-tools. En: MADELLA, M. & D. ZURRO (eds.), Plants, People and Places. Recent studies in phytolith analysis, pp. 98-106. Oxbow Books, Cambridge. https://doi.org/10.2307/jxtvh1dtr4 CAMPBELL, R. 2014. Organización y diferenciación social a través de tres comunidades de Isla Mocha (1000-1700 d.C.). Aspectos metodológicos y sus proyecciones. En: FALLABELA, F., L. SANHUEZA, L. CORNEJO & I. CORREA (eds.), Distribución Espacial en Sociedades No Aldeanas: Del Registro Arqueológico a la Interpretación Social, Serie Monográfica de la SChA n°4, pp. 2950. Sociedad Chilena de Arqueología, Santiago. CAMPBELL, R., C. ROA, A. DELGADO, C. DÁVILA, J. GAJARDO, M. LÓPEZ, I. MARTÍNEZ, G. PALMA, A. PEÑALOZA, P. ANDRADE, C. GODOY, H. INOSTROZA, C. GONZÁLEZ & M. ZÚÑIGA. 2017. Cueva de los Catalanes: nuevas investigaciones, 60 años después (Araucanía, Chile). En: GÓMEZ-OTERO, J. (comp.). X Jornadas de Arqueología de la Patagonia: libro de resúmenes. Instituto de Diversidad y Evolución Austral, Puerto Madryn.
CAMPBELL, R., H. CARRIÓN, V. FIGUEROA, A. PEÑALOZA, M. T. PLAZA & C. STERN. 2018a. Obsidianas, turquesas y metales en el sur de Chile. Perspectivas sociales a partir de su presencia y proveniencia en Isla Mocha. Chungara 50: 217-234. https://doi.org/10.4067/S0717-73562018005000501 CAMPBELL, R., C. ROA & F. SANTANA-SAGREDO. 2018b. Más sureño que los porotos: primeros fechados 14C AMS para el sitio Cueva de Los Catalanes. Boletín Sociedad Chilena de Arqueología 48: 85-59.
CARNELLI, A. L., J. P. THEURILLAT & M. MADELLA. 2004. Phytolith types and type-frequencies in subalpine-alpine plant species of the European Alps. Rev. Palaeobot. Palynol. 129: 39-65. https://doi.org/10.1016/j.revpalbo.2003.11.002
CARUSO FERMÉ, L. 2013a. Espacios interdisciplinarios en arqueobotánica: alcances y aportes para la investigación arqueológica. En: ZANGRANDO, A. F., R. BARBERENA, A. GIL, G. NEME, M. GIARDINA, L. LUNA, C. OTAOLA, S. PAULIDES, L. SALGÁN & A. TIVOLI (eds.), Tendencias Teórico-Metodológicas y Casos de Estudio en la Arqueología de la Patagonia, pp. 271279. Museo de Historia Natural de San Rafael, San Rafael.
CARUSO FERMÉ, L. 2013b. Experimentación y propiedades combustibles de especies del bosque andino-patagónico. Aportación al estudio antracológico de sitios arqueológicos. Magallania 41: 145-158.
http://dx.doi.org/10.4067/S0718-22442013000200007 CARUSO FERMÉ, L. & M. T. CIVALERO. 2014. Holocene landscape changes and wood use in Patagonia: Plant macroremains from Cerro Casa de Piedra 7. The Holocene 24: 188-197. https://doi.org/10.1177/0959683613516816 CARUSO FERMÉ, L. & M. T. CIVALERO. 2019. Fuel management in high mobility groups in Patagonia (Argentina) during the Holocene: Anthracological evidences of the sites Cerro Casa de Piedra 5 and Cerro Casa de Piedra 7. The Holocene 29: 11341144. https://doi.org/10.1177/0959683619838044 CARUSO FERMÉ, L. & A. F. ZANGRANDO. 2019. Anthracological analyses of fuel wood used by hunter-gatherers in the south coast of Tierra del Fuego during the late Holocene. Archaeol. Anthropol. Sci. 11: 2039-2048. https://doi.org/10.1007/s12520-018-0652-4 CIAMPAGNA, M. L., P. AMBRÚSTOLO & M. A. ZUBIMENDI. 2016. Estudios antracológicos en abrigos rocosos de la costa norte de Santa Cruz (Patagonia, Argentina): análisis de los sitios El Oriental y Alero 4. Intersecc. Antropol. 17: 341-352. COÑA, P. 2010[1930]. Lonco Pascual Coña. Testimonio de un cacique mapuche. Editorial Pehuén, Santiago. CRONQUIST, A. 1981. An Integrated System of Classification of Flowering Plants. Columbia University Press, New York.
EGERT, M. & M. GODOY. 2008. Semillas, cultivos y recolección al interior de una familia mapuche huilliche en Lumaco, Lanco, Región de los Ríos, Chile. Rev. Austral Cienc. Soc. 14: 51-70. https://doi.org/10.4206/rev.austral.cienc.soc.2008.n14-03 FRANK, A. 2012. Los fogones en la meseta central de Santa Cruz durante el Pleistoceno final. Magallania
40: 145-162.
http://dx.doi.org/10.4067/S0718-22442012000100009 GAJARDO, J. 2016. Análisis del material arqueológico del sitio Cueva de los Catalanes (excavación 1956). Informe Final de Práctica Profesional, Universidad de Chile, Chile.
GALANIDOU, N. 2000. Patterns in caves: foragers, horticulturists, and the use of space. J. Anthropol. Archaeol. 19: 243-275. https://doi.org/10.1006/jaar.1999.0362 GAY, C. 1865. Agricultura. Serie Historia Física y Política de Chile, Tomo Segundo. Museo de Historia Natural de Santiago, Santiago.
GAO, G., D. JIE, Y. WANG, L. LIU, H. LIU, D. LI, N. LI, J. SHI & C. LENG. 2018. Do soil phytoliths accurately represent plant communities in a temperate region? A case study of Northeast China. Veg. Hist. Archaeobot. 27: 753-765. https://doi.org/10.1007/s00334-018-0670-2 HAMMOND, H. & M. L. CIAMPAGNA. 2019. Estudio experimental de alteración térmica sobre exoesqueletos de moluscos marinos (Nacella magellanica) y sus implicancias para la interpretación de conjuntos arqueomalacológicos de concheros en Patagonia argentina. Bol. Arqueol. Exp. 13: 9-36. https://doi.org/10.15366/baexuam2018-19.13.002 HANSEN, J. 2001. Macroscopic plant remains from Mediterranean caves and rockshelters: Avenues of interpretation. Geoarchaeology 16: 401-432. https://doi.org/10.1002/gea.1010 HILGER, I. 2015. Infancia, Vida y Cultura Mapuche. Editorial Pehuén, Santiago.
JUAN I TRESSERRAS, J. 1997. Procesado y Preparación de Alimentos Vegetales para Consumo Humano. Aportaciones al estudio de fitolitos, almidones y lípidos en yacimientos arqueológicos prehistóricos y protohistóricos del cuadrante NE de la Península Ibérica. Tesis Doctoral. Universitat de Barcelona, España.
KEALHOFER, L. & D. PIPERNO. 1998. Opal phytoliths in Southeast Asian Flora. Smithson. Contrib. Bot. 88: 1-39.https://doi.org/10.5962/bhl.title.103698 LEJAY, M., M. ALEXIS, K. QUÉNÉA, F. SELLAMI & F. BON. 2016. Organic signatures of fireplaces: Experimental references for archaeological interpretations. Org. Geochem. 99: 67-77. https://doi.org/10.1016Zj.orggeochem.2016.06.002 LEROI-GOURHAN, A. & M. BRÉZILLON. 1972. Fouilles de Pincevent. Essai d'analyse ethnographique d'un habitat magdalénien (la
section 36). Éditions du Centre National de la Recherche Scientifique 15, París.
LUEBERT, F. & P. PLISCOFF. 2017. Sinopsis bioclimáticay vegetacionalde Chile. 2a Ed. Editorial Universitaria, Santiago.
LUMLEY, H. 2006. Il ya 400 000 ans: la domestication du feu, un formidable moteur d'hominisation. Comptes Rendus Palevol. 5: 149-154. https://doi.org/10.1016/j.crpv.2005.11.014 MA, J. F. 2003. Function of silicon in higher plants. En: MÜLLER, W. E. G. (ed.), Silicon Biomineralization, vol. 33: 127-147. Springer, Berlin-Heidelberg. https://doi.org/10.1007/978-3-642-55486-5_5 MADELLA, M., A. ALEXANDRE & T. BALL. 2005. International Code for phytolith nomenclature 1.0. Ann. Bot. 96: 253-260. https://doi.org/10.1093/aob/mci172 MADELLA, M. & C. LANCELOTTI. 2012. Taphonomy and phytoliths: a user manual. Quat. Int. 275: 76-83. https://doi.org/10.1016/j.quaint.2011.09.008 MADELLA, M., A. H. POWER-JONES & M. K. JONES. 1998. A simple method of extraction of opal phytoliths from sediments using a non-toxic heavy liquid. J. Archaeol. Sci. 25: 801-803. https://doi.org/10.1006/jasc.1997.0226 MARCH, R. 1992. L'utilisation du bois dans les foyers préhistoriques: une approche expérimentale. Bull. Soc. Bot. France 139: 245-253. https://doi.org/10.1080/01811789.1992.10827103 MARCH, R. 1995. Un détour vers l'histoire: l'étude des structures de combustion en archéologie. Ann. Fond. Fyssen 10: 53-68.
MARCH, R. & A. LUCQUIN. 2007. Activités liées á l'utilisation du feu et analyse des comportements: modalités fonctionnelles, modalités saisonniéres. En: BEYRIES, S. & V. VATÉ (eds.), Les civilisations du Renne d'hier et d'aujourd'hui. Approches ethnohistoriques, archéologiques et anthropologiques, Actes de XXVIIe Rencontres Internationales d'Archéologie et d'Histoire d'Antibes, 2006, pp. 1-18. Éditions APDCA, Antibes.
MARCH, R., A. BALDESSARI & E. G. GROSS. 1989. Determinación de compuestos orgánicos en estructuras de combustión arqueológicas. Mémoires du Musée de Préhistoire d'Ile de France 2: 47-58. MARCH, R., A. BALDESSARI, J. C. FERRERI, A. GRANDE, E. G. GROSS, O. MORELLO & R. RODANO. 1991. Étude des structures de combustion archéologiques d>Argentine. Bull. Soc.
Prehist. Frangaise 86: 384-392. https://doi.org/10.3406/bspf.1989.9896 MARSTON, J. M. 2014. Ratios and simple statistics in Paleoethnobotanical analysis. En: MARSTON, J. M., J. D'ALPOIM & C. WARINNER (eds.), Method and Theory in Paleoethnobotany, pp. 163-179. University Press of Colorado, Louisville. https://doi.org/10.5876/9781607323167 MASSONE, M. 2017. Fuego, Fogones y Contextos Arqueológicos de los Cazadores recolectores Tardíos en el Norte de Tierra del Fuego. Tesis Doctoral. Universidad Nacional del Centro de la Provincia de Buenos Aires, Argentina.
MASSONE, M. & M. E. SOLARI. 2017. Fogones de los cazadores-recolectores del Holoceno tardío en el norte de Tierra del Fuego: contextos y antracología. Magallania 45: 255-271.
http://dx.doi.org/10.4067/S0718-22442017000200255 MASSONE, M., A. PRIETO, D. JACKSON, G. CÁRDENAS, M. ARROYO & P. CÁRDENAS. 1998. Los cazadores tempranos y sus fogatas: una nueva historia para la cueva Tres Arroyos 1. Tierra del Fuego. Boletín de la Sociedad Chilena de Arqueología 26: 11-18.
MENGHIN, O. 1959-1960. Estudios de prehistoria araucana. Acta Praehistorica III-IV: 49-120. METCALFE, C.R. 1969. Anatomy as an aid to classifying the Cyperaceae. Am. J. Bot. 56: 782-790. https://doi.org/10.1002/j.1537-2197.1969.tb09726.x MORALES, D. V., S. MOLARES & A. LADIO. 2017. Firewood Resource Management in different landscapes in NW Patagonia. Front. Ecol. Evol. 5: 111. https://doi.org/10.3389/fevo.2017.00111 NÚÑEZ DE PINEDA Y BASCUÑÁN, F. 1863[1673]. Cautiverio Feliz, y Razón Individual de las Guerras Dilatadas del Reino de Chile. Colección de Historiadores de Chile y Documentos Relativos a la Historia Nacional, tomo III. Imprenta del Ferrocarril, Santiago.
OAKLEY, K. P. 1955. Fire as a palaeolithic tool and weapon. Proceedings of the Prehistoric Society 21: 36-48. https://doi.org/10.1017/S0079497X00017382 OLLENDORF, A. L. 1992. Toward a classification scheme of Sedge (Cyperaceae). En: RAPP, G. & S. MULHOLLAND (eds.), Phytolith Systematics. Emerging Issues, pp. 91-111. Springer Science+Business Media, New York. https://doi.org/10.1007/978-1-4899-1155-1 ORTEGA, I. M., M. I. BERGER & M. FLORES. 1993. Manual de Técnica Microhistológica. SR-CRSP y IBTA, La Paz.
OSTERRIETH, M., M. MADELLA, D. ZURRO & M. F. ÁLVAREZ. 2009. Taphonomical aspects of silica phytoliths in the loess sediments of the Argentinean Pampas. Quat. Int. 193: 70-79. https://doi.org/10.10167j.quaint.2007.09.002
PÉREZ DE MICOU, C. 1991. Fuegos, fogones y señales. Una aproximación etnoarqueológica a las estructuras de combustión en el Chubut medio. Arqueología 1: 125-150.
PERLES, C. 1977. Préhistoire du Feu. Ed. Masson, Paris.
PIPERNO, D. 1988. Phytolith Analysis. An Archaeological and Geological Perspective. Academic Press INC., San Diego.
PIPERNO, D. 2006. Phytoliths. A comprehensive guide for archaeologists and paleoecologists. Altamira Press, Oxford.
PIPERNO, D. & D. PEARSALL. 1993. Phytoliths in the reproductive structures of maize and teosinte: implications for the study of maize evolution. J. Archaeol. Sci. 20: 337-362. https://doi.org/10.1006/jasc.1993.1021
PIPERNO, D. & K. STOTHERT. 2003. Phytolith evidence for early Holocene Cucurbita domestication in southwest Ecuador. Science 299: 1054-1057. https:// doi.org/10.1126/science.1080365
QUIROZ, D. & M. SÁNCHEZ. 1997. La Isla de las Palabras Rotas. DIBAM-Centro de Investigaciones Diego Barros Arana, Santiago.
ROA, C. 2016. De la quinua mapuche a la frutilla silvestre: el aprovechamiento de recursos vegetales de importancia alimenticia en Isla Mocha (10501685 d.C.). Memoria para optar del título de Arqueóloga. Universidad de Chile, Chile.
ROA, C. 2018. Plant use in Araucanía during Late Holocene: phytolith record of Los Catalanes Cave (AD 400-1800) (Malleco Province, Chile). Tesis de Maestría. Universitat Rovira i Virgili, Tarragona.
ROA, C., D. BUSTOS, H. RAMÍREZ & R. CAMPBELL. 2018. Entre la Pampa y el Pacífico Sur. Evaluando la dispersión más austral de cultígenos en el Cono Sur americano desde la evidencia arqueobotánica y radiométrica de Isla Mocha y Cueva de los Catalanes (sur de Chile). An. Arqueol. Etnol. 73: 189- 220.
ROJAS, G. 2004. Estudios botánicos, paleoambiente y arqueología. Cerro Onas, Tres Arroyos, Tierra del Fuego. Chungara 36: 381-386. http://dx.doi.org/10.4067/S0717-73562004000300040
ROSEN, A. M. 1992. Preliminary identification of silica skeletons from Near Eastern archaeological
sites: an anatomical approach. En: RAPP, G. & S. MULHOLLAND (eds.), Phytolith Systematics. Emerging Issues, pp. 129-139. Springer Science+Business Media, New York. https://doi.org/10.1007/978-1-4899-1155-1 SHENNAN, S. 1992. Arqueología Cuantitativa. Editorial Crítica, Barcelona. https://doi.org/10.1016/C2009-0-21716-8
SHILLITO, L. M. 2011. Taphonomic observations of archaeological wheat phytoliths from Neolithic Qatalhoyük, Turkey, and the use of conjoined phytolith size as an indicator of water availability. Archaeometry 53: 631-641. https://doi.org/10.nn/j.1475-4754.2010.00582.x SHILLITO, L. M. & W. MATTHEWS. 2013. Geoarchaeological investigations of midden-formation processes in the Early to Late Ceramic Neolithic levels at Qatalhoyük, Turkey ca. 85508370 cal BP. Geoarchaeology 28: 25-49. https://doi.org/10.1002/gea.21427 SMITH, A. 2014. The use of multivariate statistics within Archaeobotany. En: MARSTON, J. M., J. D'ALPOIM & C. WARINNER (eds.), Method and Theory in Paleoethnobotany, pp. 181-204. University Press of Colorado, Colorado. https://doi.org/10.5876/9781607323167 SOLARI, M. E. 1991. Estudio antracológico del sitio Punta Baja 1 (Mar de Otway). An. Inst. Patagon. 19: 115-120.
SOLARI, M. E. 1992. Anthracologie et ethnoarchéologie dans l'archipel du cap Horn (Chili). Actualités Botaniques 139: 407-419. https://doi.org/10.1080/01811789.1992.10827117 SOLARI, M. E. 1993. L'homme et le bois en Patagonie et Terre de Feu au cours des six derniers millénaires: recherches anthracologiques au Chili et en Argentine. Thése de Doctorat. Université de Montpellier II, France. SOLARI, M. E. 1994. Estudio antracológico del archipiélago del Cabo de Hornos y Seno Grandi. An. Inst. Patagon. 22: 137-148.
SOLARI, M. E. 2000. Antracología, modo de empleo: en torno a paisajes, maderas y fogones. Rev. Austral Cienc. Soc. 4: 167-174.
https://doi.org/10.4206/rev.austral.cienc.soc.2000.n4-SOLARI, M. E. 2003. Análisis antracológico del Locus 1. Magallania 31: 327-335.
SOLARI, M. E. 2009. Análisis antracológico de una columna estratigráfica del alero cerro Castillo (Magallanes, Chile). Magallania 37: 157-162. https://doi.org/10.4067/S0718-22442009000100012
SOLARI, M. E., V. BERNARD, D. LEGOUPIL, H. RICHARD, P. ROIRON & P. SCHOELLAMMER. 2002. Palaeoenvironmental approach of Ponsonby archaeological site (Riesco Island, Chilean Patagonia). En: THIBAULT, S. (ed.), Charcoal analysis. Methodological Approaches, palaeoecological results and wood uses, B.A.R. International Series 1063, pp. 179-185. https://doi.org/10.30861/9781841714431 SOLARI, M. E. & C. A. LEHNEBACH. 2004. Pensando la antracología para el centro-sur del Chile: sitios arqueológicos y bosque en el Lago Calafquén. Chungara 36: 373-380.
http://dx.doi.org/10.4067/S0717-73562004000300039 THÉRY-PARISSOT, I., 1998. Économie du combustible au Paléolithique. Anthracologie, Expérimentation, Taphonomie. Thése de Doctorat. Université Paris 1 Panthéon-Sorbonne, Paris.
TORRES, L., O. PARRA, A. ARANEDA, R. URRUTIA, F. CRUCES & L. CHIRINOS. 2008. Vegetational and climatic history during the late Holocene in lake Laja basin (central Chile) inferred from sedimentary pollen record. Rev. Palaeobot. Palynol. 149: 18-28. https://doi.org/10.1016/j.revpalbo.2007.10.001 TSARTSIDOU, G., S. LEV-YADUN, R. M. ALBERT, A. MILLER-ROSEN, N. EFSTRATIOU & S. WEINER. 2007. The phytolith archaeological record: strengths and weaknesses evaluated based on a quantitative modern reference collection from Greece. J. Archaeol. Sci. 34: 1262-1275. https://doi.org/10.1016/jjas.2006.10.017 TWISS, P., E. SUESS & R. M. SMITH. 1969. Morphological Classification of Grass Phytoliths. Soil Sci. Soc. Am. J. 33: 109-115. https://doi.oig/102136/sssai1969.03615995003300010030x
VALLVERDÚ, J., S. ALONSO, A. BARGALLÓ, R. BARTROLÍ, G. CAMPENY, A. CARRANCHO, I. EXPÓSITO, M. FONTANALS, J. GABUCIO, B. GÓMEZ, J. M. PRATS, P. SAÑUDO, P. SOLÉ, J. VILALTA & E. CARBONELL. 2012. Combustion structures of archaeological level O and mousterian activity areas with use of fire at the Abric Romaní rockshelter (NE Iberian Peninsula). Quat. Int. 247: 313-324. https://doi.org/10.1016Zj. quaint.2010.12.012
WEINER, S. 2010. Microarchaeology. Beyond the Visible Archaeological Record. Cambridge University Press, New York. https://doi.org/10.1017/CBO9780511811210 ZURRO, D. 2006. El análisis de fitolitos y su papel en el estudio del consumo de recursos vegetales en la prehistoria: bases para una propuesta metodológica materialista. Trab. Prehist. 63: 35-54. https://doi.org/10.3989/tp.2006.v63.i2.16 ZURRO, D. 2011. Ni carne ni pescado (consumo de recursos vegetales en la Prehistoria): Análisis de la variabilidad de los conjuntos fitolitológicos en contextos cazadores- recolectores. Tesis Doctoral. Universitat Autónoma de Barcelona, España. ZURRO, D. 2018. One, two, three phytoliths: assessing the minimum phytolith sum for archaeological studies. Archaeol. Anthropol. Sci. 10: 1673-1691. https://doi.org/10.1007/s12520-017-0479-4
ZURRO, D., M. MADELLA, I. BRIZ & A. VILA. 2009. Variability of the phytolith record in fisher-hunter-gatherer sites: An example from the Yamana society (Beagle Channel, Tierra del Fuego, Argentina). Quat. Int. 193: 184-191. https://doi.org/10.1016/j.quaint.2007.11.007

