











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink1. INTRODUCCIÓN
Como respuesta al cambio climático global el sector forestal ha ido ampliando el foco del manejo desde la orientación exclusiva de la producción maderera hacia una visión que incorpore también los servicios ambientales (Bravo et al. 2008). Esto conlleva la necesidad de conocer la tasa y los patrones de partición de la biomasa en los distintos compartimentos del árbol para luego estimar la demanda de nutrientes en rodales de especies de rápido crecimiento (Adegbidi et al. 2002) y evaluar la sostenibilidad de estos sistemas. En los últimos años, el cultivo del Pinus taeda en Misiones, ha sido acompañado de una fuerte expansión del cultivo del pino híbrido, entre otras características por su mejor forma, sus altas tasas de crecimiento y menor tamaño de ramas, entre las principales características, por lo que también resulta necesario conocer el aumento de la productividad de esta especie en relación a las tradicionales para la producción maderera en la principal región forestal de Argentina.
En los bosques implantados están cobrando importancia aquellos desarrollados a partir de híbridos interespecíficos. En Australia durante la década de 1960 se iniciaron trabajos de hibridación entre Pinus elliottii var. elliottii (Engelm) x Pinus caribaea var. elliottii (Morelet) (Nickles 1992). Como resultado de estos cruzamientos se obtuvieron materiales con un comportamiento superior al de los parentales, despertando el interés en otras regiones forestales del mundo y Sudamérica (Nickles 1991, Dieters et al. 2006). Crechi et al. (2004) evaluaron la respuesta del pino hibrido y a la oportunidad e intensidad de raleos, encontrando diferencias significativas sobre el incremento y la producción total.
Cuando se quiere predecir más de un componente de la biomasa aérea de un árbol, es deseable que el procedimiento utilizado contemple las propiedades de aditividad (Borders 1989, Parresol 1999). Esto significa que la suma de las predicciones para cada componente corresponde a la predicción para la biomasa total del árbol (Carvalho et al., 2003, Huiquan et al., 2004). Zellner (1962) describió el método de estimación de regresiones aparentemente no relacionadas conocido como SUR (seemingly unrelated regressions). El mismo permite estimar los parámetros de un conjunto de ecuaciones de regresión de forma asintóticamente más eficiente que los obtenidos por la ecuación de mínimos cuadrados (Borders, 1989; Parresol, 1999). Este procedimiento ha sido empleado en econometría (Zivot y Wang, 2006) y en los últimos años, ha comenzado a aplicarse en los modelos de predicción de biomasa de árbol individual, pero teniendo en cuenta la correlación entre arboles dentro de la parcela (Rose, 2001). Russell et al., 2009 desarrollaron estimaciones SUR en parcelas de árbol individual de 6 años de edad para determinar la biomasa aérea de Pinus taeda y el efecto de la densidad inicial de plantación. Schneider et al. (2008) calibraron las relaciones alométricas de Pinus banksiana Lamb. usando este tipo de estimaciones e incorporando variables de calidad de sitio que mejoraron las predicciones cuando se trabaja a escala regional. Sanquetta et al (2015) en Brasil , trabajando con Pinus elliottii, desarrollaron ecuaciones SUR para fracciones de biomasa de árboles individuales y encontraron mejores ajustes que estimando las componentes por separado.
Este trabajo tiene como objetivo estimar las componentes de modelos de biomasa aérea de pinos híbridos Pinus elliottii var. elliottii x Pinus caribaea var hondurensis para la región agroecológica Misiones Norte de Argentina (Papadakis, 1974) aplicando modelos aditivos ajustados con la metodología SUR y contrastarlos con los modelos ajustados por estimadores mínimos cuadrados ordinarios MCO.
2. MÉTODOS
Área de estudio
La zona de donde se obtuvieron las muestras corresponde a la región fitogeográfica de la provincia Paranaense (Dominio Amazónico, Cabrera 1976), distrito de las selvas (Rodríguez et al., 2005), que se corresponde con la zona agroecológica Misiones Norte (Figura 1). Los suelos de la región pertenecen al gran grupo de los Kandiudultes (US Soil Taxonomy) conocidos localmente como tierra colorada o suelo rojo profundo (Mancini et al. 1964 y Ligier et al. 1990). Los mismos son derivados de la meteorización de la roca basáltica, siendo arcillosos en todo su perfil (INTA, 1972 - 1997). El clima según la clasificación de Köppen es de tipo Cfa (templado húmedo sin estación seca, macrotérmico, constantemente húmedo y subtropical) (Rodríguez et al., 2005). La precipitación media durante el período 1971-2011, fue de 1.885 mm en Iguazú, ciudad localizada al norte de la región considerada. La temperatura media anual en el mismo período fue de 21,6 ºC (Olinuck, 2012).
Base de datos
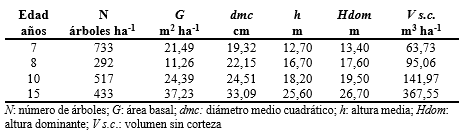
Se constituyó con 24 ejemplares cortados en cuatro parcelas de muestreo establecidas en rodales de dos empresas de la región, Lipsia SA y Trumpp Hnos. (Figura 1). Los ejemplares tenían distintas edades de manera de representar el rango de las plantaciones comerciales existentes. Tres rodales tenían un manejo forestal intensivo con raleos comerciales tempranos y poda hasta 8 m de altura y pertenecían a la empresa Lipsia SA, ubicada en el departamento Iguazú y un cuarto con manejo silvopastoril, con aplicación de raleos precomerciales, podas y un raleo comercial, estaba ubicado en la empresa Trumpp Hnos del Departamento Montecarlo de la provincia de Misiones (Tabla 1). El muestreo se circunscribió a esos cuatro rodales, ya que su cultivo en la región aun no está suficientemente extendido y no hay mayor diversidad de sitios, clases de edades y regímenes silvícolas.
En las parcelas de muestreo se procedió a la medición del diámetro a 1,3 m (d) de todos los ejemplares. En cada una de ellas los árboles fueron ordenados por categoría diamétrica y divididos en tres clases: “pequeños”, “intermedios” y “grandes” (Köhl et al., 2006), siendo los límites de dichas clases variables para cada lote analizado en función de su edad y manejo”. De cada clase diamétrica se seleccionaron al azar dos ejemplares, cortándose seis árboles por parcela para la determinación de biomasa aérea. De los árboles cortados se midieron las alturas totales con las se construyeron relaciones hipsométricas para cada parcela, que permitieron estimar la altura de cada ejemplar, de modo de calcular la altura media (h) y la altura dominante (Hdom). Esta información permitió caracterizar los parámetros dasométricos de las parcelas de donde se obtuvieron los árboles muestra (Tabla 1).
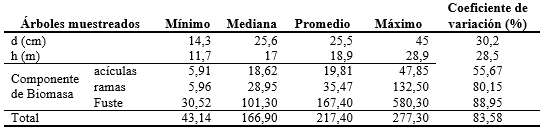
De cada ejemplar cortado se procedió a determinar el peso fresco total de acículas verdes y el de ramas. De estas fracciones se tomaron alícuotas que fueron llevadas a estufa a una temperatura de 70 ºC hasta peso constante, aplicando luego los coeficientes obtenidos a los valores de peso verde para estimar las biomasas de acículas y ramas. El fuste del árbol fue trozado en largos comerciales hasta los 5 cm de diámetro en punta fina y en cada extremo de las trozas se tomaron dos diámetros en forma perpendicular con y sin corteza y se cortaron discos. El volumen de cada rollizo fue determinado mediante la fórmula de Smalian. De los discos, desde el centro hacia la periferia en ambos sentidos, se obtuvieron probetas de 2 x 2 cm, las cuales fueron llevadas a estufa a una temperatura de 103 ºC+/-2 ºC hasta peso constante para estimar densidad básica. Ésta se determinó mediante el método de desplazamiento volumétrico. El valor de densidad básica promedio obtenido de los discos de cada rollizo se aplicó luego al volumen sin corteza de los mismos para estimar su peso seco y de los árboles (Adegbidi et al., 2002, Pinazo et al., 2007). El volumen de corteza del fuste no se consideró en estos cálculos. Los valores máximos, mínimos y promedios de las variables descriptoras de las parcelas y de los ejemplares cortados se presentan en la Tabla 2.
Análisis estadístico. Los modelos ajustados de forma simultánea para cada componente fueron previamente seleccionados entre los reportados comúnmente en la literatura (Burkhart, 2012) (Tabla 3). Las variables explicatorias fueron d y h. Las variables dependientes fueron: biomasa de acículas (BAc), biomasa de ramas (BR), biomasa del fuste (BF) y biomasa total (BT) todas expresadas en kilogramos. Se aplicaron transformaciones logarítmicas de las variables dependientes e independientes para lograr la homogeneidad de las varianzas y linealizar los modelos. Una vez realizadas las estimaciones se seleccionó aquel que mejor ajustaba para cada componente de biomasa en función de la significancia de los estimadores, los valores de R2 (coeficiente de determinación) y del CME (cuadrado medio del error).
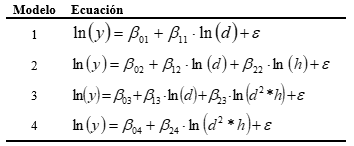
En una segunda etapa se procedió al ajuste simultáneo de todas las componentes de biomasa bajo un sistema de ecuaciones que puede representarse en forma matricial según [1]
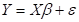 [1]
[1]
Donde:
Y = la matriz de p variables dependientes,
X = la matriz de incidencia para cada variable independiente,
β = vector de efectos a estimar,
ε = el vector de errores aleatorios y se asume que su distribución es Normal Multivariada 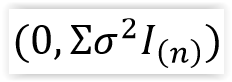 .
.
Zellner (1962) formuló las regresiones aparentemente no relacionadas (SUR), como p ecuaciones de regresión, con posibles correlaciones entre los términos de error de las mismas. El modelo SUR se explicitó según el sistema matricial [2]
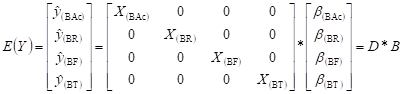 [2]
[2]
El vector de coeficientes se estimó utilizando la fórmula [3]
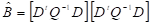 [3]
[3]
Donde Q es la matriz de ponderación basada en las covarianzas de las Y variables, según la forma [4]
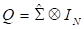 [4]
[4]
Los elementos de la matriz 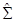 de la ecuación [4] se calcularon como [5]
de la ecuación [4] se calcularon como [5]
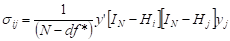 [5]
[5]
Donde: 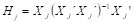 .
.
Como puede verse los modelos SUR son una aplicación particular de los modelos lineales generalizados.
Ajuste por transformación. Por último se estimó un factor de corrección según la ecuación [6] con el objetivo de eliminar la asimetría en la distribución que se produce en el proceso de transformación inversa, que se da al emplear la transformación logarítmica (Meyer 1941, Baskerville 1972).
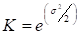 [6]
[6]
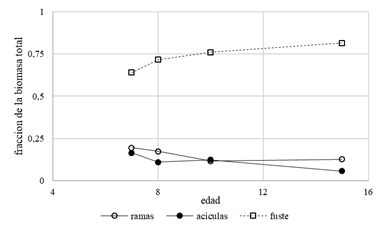
Donde: K= coeficiente de corrección de la discrepancia logarítmica y σ2: varianza.
Los cálculos se realizaron en un entorno R (R Development Core Team 2012), aplicando las funciones de los paquetes systemfit (Arne y Hamann 2007) y nlme (Pinheiro et al., 2011).
3. RESULTADOS
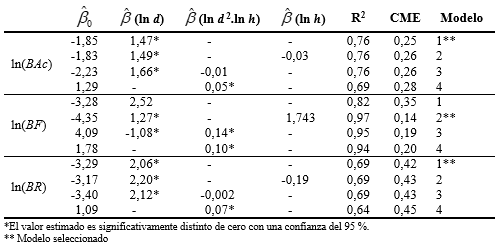
En el ajuste de los modelos independientes para cada componente de biomasa (Tabla 4), el modelo 1 fue el que mejor ajustó para biomasa de acículas (BAc) y biomasa de ramas (BR) dado que en ambos casos presentó el menor CME y el mayor R2, siendo la contribución de estas componentes a la biomasa total del 14 y 12 % respectivamente. Para biomasa del fuste (BF), siguiendo los mismos criterios se optó por el modelo 2 que además del d incluye la altura. Para dicha componente la contribución a la biomasa total es del 74 %.
Tabla 4 Valores estimados de los distintos parámetros de los modelos propuestos y sus coeficientes de ajuste (BAc = biomasa de acículas, BF = biomasa del fuste, BR = biomasa de ramas)
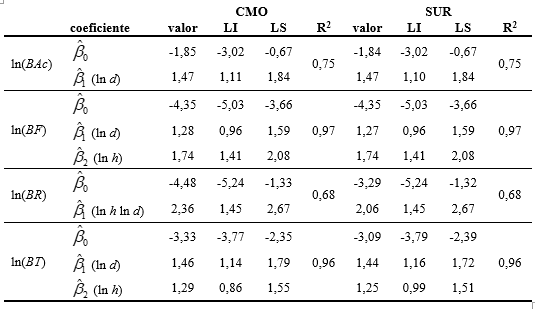
Bajo el ajuste simultáneo de los modelos seleccionados los valores de los parámetros resultaron muy similares en todas las fracciones al comparar los estimadores SUR y CMO (cuadro 5), excepto para biomasa de ramas donde los estimadores SUR tiene valores de ordenada al origen y de pendiente menores a los valores ajustados con estimadores CMO. También se registró una reducción leve en los intervalos de confianza al utilizar la metodología SUR respecto a la CMO (Tabla 5), haciéndolos más informativos.
Puede verificarse que en el ajuste de biomasa de ramas el 41% de las observaciones presentaron errores por debajo del 10 % cuando se aplicó el método SUR, en tanto que solo el 8 % de las observaciones presentaron errores de menos del 10 % cuando se ajustó por el método de CMO. En este último caso, el 58 % de las observaciones presentaron errores de más del 20 %. También se vio un cambio en los valores de correlación entre los estimadores de las componentes de varianza, mientras las mismas se asumen cero cuando se estiman los parámetros con el método CMO. Para los estimadores SUR se encontró que la correlación entre la ordenada al origen de la biomasa de ramas y acículas fue del 17 %, y de un 5 % para la pendiente. La matriz de correlaciones entre los residuales de los modelos CMO de las distintas componentes fue:
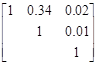
De derecha a izquierda las columnas corresponden a biomasa de acículas, ramas y biomasa del fuste. La prueba estadística para determinar si Ho: 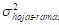 =0 indicó correlación significativa entre biomasa de ramas y acículas, consecuentemente el método SUR provee estimadores más eficientes que los obtenidos por CMO. Esto se ve reflejado en un cambio de los valores estimados y en el comportamiento de los errores de estimación.
=0 indicó correlación significativa entre biomasa de ramas y acículas, consecuentemente el método SUR provee estimadores más eficientes que los obtenidos por CMO. Esto se ve reflejado en un cambio de los valores estimados y en el comportamiento de los errores de estimación.
Comparando la relación entre los valores observados y ajustados por ambos métodos, para el caso de biomasa de ramas los predichos del modelo SUR adopta valores mayores a los del modelo CMO para toda la serie. Esta diferencia tiende a ser levemente mayor al d. (Figura 2). Estos valores fueron ajustados por coeficientes K para eliminar la asimetría en la distribución que se produce en el proceso de transformación inversa, cuyo valor para biomasa de ramas fue de 1,1.
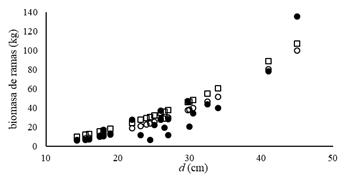
Figura 2 Valores observados (●), estimados CMO (○) y estimados SUR (□) de biomasa de ramas en función del diámetro a la altura de pecho (d).
El modelo de ajuste para biomasa del fuste que es la principal contribuyente a la biomasa total no mostró diferencias entre ambos métodos de estimación (Figura 3).
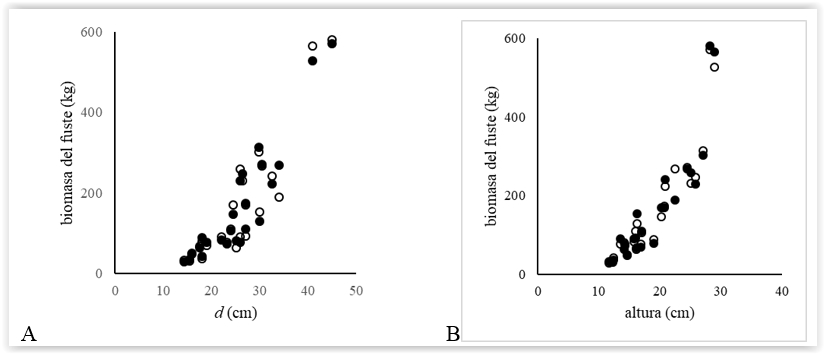
Figura 3 A.- Valores observados (○) y estimados (●) de biomasa del fuste en función del diámetro a la altura de pecho (d). B.- Valores observados (○) y estimados (●) de biomasa del fuste en función de la altura
La contribución de la biomasa del fuste aun a edades tempranas del rodal, fue mayor al 60 %, siendo del 82 % a los 15 años. La fracción de ramas parecería estabilizarse en 11 % a los 10 años, en tanto que la biomasa de acículas varió del 16 % en árboles de 7 años a 6 % en los de 15 años (Figura 4).
4. DISCUSIÓN Y CONCLUSIONES
Se obtuvieron modelos aditivos de predicción de biomasa aérea de pinos híbridos útiles para estimar dichas componentes en función del d y la altura. Estos pueden ser utilizados como herramientas de cálculo siendo promisorias para la investigación sobre modelos y sistemas locales de simulación de crecimiento y producción. En concordancia con lo encontrado por Russell et al. (2009) para Pinus taeda, las estimaciones SUR mejoraron los resultados de las estimaciones CMO En el presente trabajo se obtuvo una mejora en el caso de las ramas. Se verifica que todas las funciones obtenidas resultan ser biológicamente razonables, acorde a lo recomendado por Carvalho y Parresol (2003), respecto al mantenimiento de la independencia y aditividad de las componentes. Las estimaciones de los parámetros resultan más eficientes (Schneider et al., 2008).
Schneider et al. (2008) trabajando con Pinus banksiana reportaron modelos aditivos considerando la proporcionalidad del área basal y el follaje en relación a la densidad del rodal. En el presente trabajo, las características de los rodales muestreados fueron de baja densidad, con arboles creciendo prácticamente sin competencia.
Las relaciones alométricas alcanzaron ajustes satisfactorios para biomasa del fuste, en cambio los modelos correspondientes biomasa de acículas y biomasa de ramas requiere que los resultados obtenidos deban ser utilizados con precaución debido a la variabilidad presente. Los valores de R2 de las distintas componentes de biomasa son semejantes a los publicados por Sanquetta et al. (2000) en Pinus taeda y los coeficientes del modelo están dentro del rango, reflejando las diferencias genéticas y ambientales
En concordancia con lo presentado por Fassola et al (2012), el pino hibrido mejora la asignación de biomasa a fuste en detrimento de las ramas en relación al Pinus taeda.
Dado que la base de datos es reducida estos modelos son sólo factibles de emplear para cuantificar la biomasa aérea en forma preliminar en plantaciones híbridas (F2) a nivel local. El incremento futuro del área de plantación, como el seguimiento y análisis destructivo de ensayos instalados, deberán subsanar las restricciones de estos modelos, permitiendo formular hipótesis acerca de la partición de biomasa. La adición de variables de caracterización del sitio, tales como densidad, índice de sitio, temperatura y humedad, podrían ser evaluadas para una formulación de ecuaciones a nivel regional.
Las estimaciones SUR resultan ser el mejor método para las predicciones de las fracciones de biomasa por la compatibilidad de las ecuaciones y la consistencia de sus coeficientes.

