











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink1. INTRODUCTION
In tropical and subtropical forests, photosynthetic active radiation influences the development of understory vegetation and its spatial distribution (Chazdon, 1986; Campanello et al., 2007). Much of the variability in tree growth rates, both within and among species, can be accounted for the assumption that growth is proportional to light interception (King et al., 2005). Most Neotropical understory palm species are shade tolerant (Hodel, 1992), however, they can also take advantage of small forest gaps (Nakazono et al., 2001). Human activities can modify the physical environment of an ecosystem by altering structural features. Particularly, in forests under harvesting regimes, the extraction of the overstory vegetation creates canopy openness, which generates an increase in the light conditions in the understory (Chen et al., 1999). These forest changes affect regeneration patterns and development of the species, especially the exploited ones, compromising the economic yield and the ecological sustainability of the forest.
Euterpe edulis Mart. (Arecaceae) is a monopodial arborescent palm endemic to the Atlantic Forest of Brazil, Paraguay and Argentina (Henderson et al., 1995). It is harvested for its heart palm that constitutes an economically important forest product (Galetti and Fernández, 1998). The palm heart extraction implies the death of the individual. Intensive harvest of this palm specie has caused a large reduction of its populations and local extinction in several areas (Henderson et al., 1995; Galetti and Fernández, 1998). Sustainable management of this palm tree must be designed taking into consideration individuals and population growth rates in relation to environmental conditions.
Tomlinson (1990) suggested that characterized phases of development are largely arbitrary but recognized that it is useful to describe the vegetative and reproductive growth of the palm in discrete phases. Seedling phase lasts until the seed endosperm is exhausted. In the sapling phase, there is a gradual increase in axis diameter. Axis progressively increases in size and serves as a root-generating region to develop an efficient anchoring and absorbing system. The next phase begins when the palm starts increasing in height. The last two phases is what Tomlinson (1990) considered the establishment step. Consistently, Alves et al. (2004) studied E. edulis allometry and explained that the relationship between stem diameter and height is not the same along the palm life. They suggested that this palm tree is able to grow to tree stature by initially overbuilding the stem at ground level. Later, there is a shift to a greater stem increment in height, and lower increment in diameter. Rich (1986) suggested that diameter growth is a product of more stiffness and strength of stem tissue. Reis et al. (2000) indicated that the increment of stem diameter measured at 1.3 meters above ground (DBH) decreases with DBH, average annual DBH growth changes from 0.04 cm on palms with DBH larger than 14 cm to 1.15 cm on palms with a DBH of eight cm.
Given that the establishment phase of palms seems to be highly significant for the entire life of the individual, studies of E. edulis’ development have centered mainly on seedling and sapling phases (Favreto et al., 2010; Gatti et al., 2011). Gatti et al. (2011) suggested that saplings can adjust their morphology and development to changes in environmental conditions, mainly light. One strategy of smaller palm trees to face low radiation is to grow proportionally more in height than in diameter, to decrease leaf production rate, and to increase leaf longevity. This determines the ontogenetic development of the palm, which is slower under low light radiation levels than under relatively high light radiation levels. On the other hand, to survive under moderately high radiation levels, individuals invest proportionally more biomass on roots than on other tissues (Gatti, 2005), decrease leaf size and increase the relationship between the surface of conductive and demanding water tissues (stem versus leaf area) (Gatti et al., 2014).
It has been observed that density has an important effect on E. edulis regeneration at a population level. Silva Matos et al. (1999) demonstrated that high seedling and adult densities of this species restrict seedling growth and survival. Fantini and Guries (2007) suggested that seedling and sapling growth rates decrease with density, in pure as well as in mixed forest. These studies inferred that the time required for individuals of E. edulis to reach a certain size may vary depending on environmental conditions.
Most tree harvest sustainable plans are based on plant size, such as DBH. But, despite of all the precedent evidences, most of the forest managers seldom consider the effect on environmental conditions on the time to reach harvest size, which is important from an ecological and economical points of view.
Argentinean law recommends to harvest palm trees with DBH equal or larger than 10 cm. Chediack (2008a) suggested that sustainable management should include includes harvesting cycles of 13-20 years, felling the palms from September to December to avoid Curculionidae infections, and leaving a set of different reproductive palms to allow genetic diversity.
Our goal was to study the growth rate and development of E. edulis palms in protected versus harvested forests. We tested the hypothesis that these two forest types affect the development rate and form of growth of the remaining individuals in the population. This could be due to changes in the forest structure and increment in canopy openness as a result of removing canopy and mid-canopy palms. Consequently, we expected that palms of similar size will differ in age when comparing forest with and without palm harvest.
2. MATERIALS AND METHODS
Study site: This study was conducted in the semi deciduous Atlantic Forest of northeastern Argentina (Oliveira Filho and Fontes, 2000) in Misiones Province. The Atlantic forest is a “hotspot of biodiversity” because of its exceptional concentration of endemic species and high rate of deforestation (Myers et al., 2000). Argentina preserves one of the most important remaining of this continuous forest, covering approximately 920,000 ha (SAyDS, Ministerio de Salud, 2005).
The climate is subtropical humid without a dry season. Average annual precipitation is 1949.5 mm. Mean annual temperature is 21 °C (SMN, 2018). In winter, there are days with subzero temperatures at dawn (Gatti et al., 2008).
Study sites were located in harvested and protected forest areas in northern Misiones Province, Argentina (around 54°13’ W and 25°4’ S). The protected, unlogged area is within Iguazu National Park (PNI) that has an area of 67720 ha. The harvested area is located in a private property next to the park, named “Establecimiento Forestal San Jorge” (ESJ). It had 6,800 ha of E. edulis dominated forest, and palm trees had been harvested eleven years prior to establishing the study (ESJ managers, pers. comm.). Forest structure, basal area, tree palm density, species diversity and number of endangered species from both sites were described by Chediack (2008b). E. edulis density in Misiones province depends on forest management. Sometimes owners cut their palms as soon as the palm reaches the legal DBH limit, or when they need cash, therefore some forests do not have any harvestable palm trees. Some landowners leave 60 adult palms per hectare as recommended by Reis et al. (2000), and others leave up to 200 adult palms per hectare (Chediack, 2008b).
Study methods
E. edulis population was studied in three permanent plots of 1 ha each. Permanent plots 1 (PP1) and 2 (PP2) were established in PNI. The third plot was established in ESJ. Plots were measured annually for three consecutive years (from 1997 to 2000) following the methods of the Smithsonian Institution Diversity Program (Dallmeier, 1992). Each permanent plot consisted of 25 subplots of 20 x 20 m. Within each subplot, every palm tree with a woody stem was measured and identified with a numbered metallic tag. Seedlings and saplings were tagged in 25 small plots of 2x2 m placed in the center of each subplot. Seedling and sapling census were carried out annually before the ripe fruits fell, that is, before the beginning of a new seed contribution.
Individuals were classified into five size categories based on Tomlinson (1990) criterion and adapted accordingly the system used by local residents and logging companies:
-
1- Seedling: Small palm tree that has just germinated and depends on seed reserves.
2- Sapling: Palm tree that no longer depends on seed reserves, it has a non-woody axis.
3- Immature: Palm tree that has a woody stem with leaf scars visible at its base. It is shorter than 1.3 m (from the base of the palm to the base of the folded leaves), so DBH is not measurable.
4- Category 4: Individuals with a woody stem at 1.3 m, and with DBH less than 10 cm.
5- Category 5: Palms with DBH larger than 10 cm.
For each category the following measurements were recorded:
Seedlings: Were counted but not measured. We arbitrary consider this phase to include the first year of life.
Saplings: Height (h) and axis diameter (D) were recorded, D was measured at the palm tree base that is at ground level.
Immature and categories 4 and 5: Total height (H) was measured from the base of the stem to the top of the tree. Not having calipers, perimeter at the stem base (PAB) was only measured on non-reproductive individuals. The PAB of reproductive palms was not measured to avoid damaging the numerous seedlings surrounding the putative parent palm.
Categories 4 and 5: Stem perimeter at 1.3m from the ground (PBH) was measured on all individuals. PBH was used to estimate individual basal areas (ba) of palm trees in category 5.
Data analysis
Basal Area: BA is the sum of ba per ha (BA=∑ ba). BA represents crowding as an independent variable and was estimated because peasants and logging companies use it as a reference to plan estimate harvests. The law allows cutting down palm trees with DBH>10 cm, therefore density of category 5 and BA changes after harvest.
Annual individual growth in size: or current annual increment (CAI), was calculated by comparing measurements averaged from two successive years.
Simple linear regression was used to model the relation between CAI as dependent variable and size measured at the first year as independent variable. For each regression, parameters slope, a (intercept) and r (regression coefficient) were tested to establish if they were significantly different from zero. Regression’s parameters from each plot were compared using Tukey and Student tests. Logarithmic regressions between PBH and height were performed, and the regression parameters from each site were compared. When regression parameters from PP1 and PP2 were not significantly different, data were pooled as it was the case in the NP (National Park). To model the dependency of CAI in palm size and BA, nonlinear multiple regressions were performed between CAI as dependent variable, and size and BA as independent variables.
Age determination: E. edulis did not present any single that could be used to predict size increments. Leaves do not leave stem scars that could be used along with leaf production rate to determine age, as it is the case with other arborescent palm species (Lugo and Rivera Battle, 1987; Renninger et al., 2013). This forced us to use three different plant development attributes to estimate palm age. Annual increment in axis diameter for saplings, palm height for the immature, and PBH for categories 4 and 5.
The palm aged is the computed counting the amount of annual increments needed to attain the size that indicates that the palm has develop in the next category size.The age at which a palm tree sapling becomes immature was called S age. Saplings convert into immature when they develop a woody axis. The age at which a palm tree develop from immature into category 4 was denominated I, which is the years that a palm spends as immature plus S. The age at which a palm moves into category 5 is C (Fig.1).

For modeling this palm tree age, constant values of BA were used. As BA changes during tree life, especially in harvested areas where the forest is under a recovery process, we contemplated situations where BA could be consider unchangeable. Three situations with constant BA were simulated: 1) Without Category 5 palms or BA=0; 2) With 60 category 5 palms per hectare; 3) With 495 category 5 palms per hectare. The last is the average palm density found by Chediack (2008b) in non-harvested forests (PNI). We consider palm tree density constant in the park because its population is regulated by density-dependence, and population growth rate rounded one (Chediack, 2014).
Sapling age was defined using saplings axis CAI in diameter, a diameter was defined for each successive age (m) as:
Diameter (m) = initial diameter + CAI diam (1) +…..+ CAI diam (m)
This formula was used for all diameters between initial diameter and Dmax (maximum axis diameter that a sapling could attain in certain AB).
For each plot, Dmax was estimated using the average of the palm axis diameter in the survey prior to the one in which the sapling develop into an immature palm. Since only 3 sites were studied, there is not enough data to establish a statistical relation. However, assuming that with three points a limit could be offered, relations that best fit the data were used to explain the dependence of Dmax for BA.
S age is the amount of years of the sapling period plus 1, the year that we assumed they last as seedling.
Age of immature palm trees was estimated using height CAI. In each plot, immature palms minimum and maximum height was estimated. Minimum height (Hmin) was the average height that palms have the first year recorded as immature. The maximum height was the average height of the palms right before they change into category 4. Height for immature was estimated for each successive age (m) based on the following equation:
Height (i) = Hmin + CAI height (1) +…..+ CAI height (i)
Where Height (i) is composed between Hmin and Hmax.
I, immature’s age at which palm becomes category 4 and it was estimated as: I=S+the years needed to reach Hmax.
Age estimation for palms in categories 4 and 5 is equal to S plus I, plus the time to reach certain PBH (This last component is calculated using CAI in PBH). Minimum PBH (PBHmin) is the stem perimeter when the palm enters into category 4. PBHmin and BA relationship were set using the regression that best fit the relation.
C is the age at which individuals in category 4 enter into category 5.
C= S+I+ the time needed to reach a DBH of 10 cm.
The relation between C and BA was estimated with the regression analysis that best fit data.
Regressions were considered only if residuals were normal (Anderson-Darling or Kruskal-Wallis tests were performed) and if the p-value was less than 0.001.
3. RESULTS
Palm tree Basal Area: The estimated BA was 7.19 m2 in PP1, 5.47 m2 in PP2 and 0.82 m2 in ESJ.
Annual individual growth: Palm trees that grow in ESJ have larger CAI in axis sapling, in immature height, and in PAB and PBH of categories 4 and 5.
- Saplings’ growth in diameter and height:
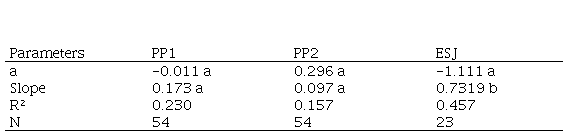
CAI in diameter was associated with individual size, the greater the axis of the sapling, the greater the diameter increment. The slope in ESJ regression was almost five times higher than in the NP. It means that saplings with the same axis diameter grew more in ESJ than in NP (Table 1). Axis diameters measured in NP ranged between 1.2 mm and 26.8 mm, and in ESJ between 2 and 19.7 mm.
Table 1. Parameters of the regression between axis annual increments in diameter and axis diameters (D) for Saplings. Intercepts were not significant different form cero. Tukey and Student tests show that slopes in PP1 and PP2 were similar to each other and they differed from ESJ’s slope (PP1-ESJ: q=13.1, P<0.001; PP2- ESJ q=15.9, P<0.001; NP-ESJ: q=4.2, P<0.05). PP1 and PP2, permanent plots in the Iguazu National Park. ESJ, harvested area. Different letters indicate significant differences
Saplings’ CAI in height was related to axis height (PP1 R² =0.4128, N=54, P<0.001; PP2 R² =0.079, N=54, P<0.001; ESJ R² =0.2158, N=23, P<0.001). But, there were not significant differences for CAI in height between sites (F=1.79, P=0.18). Average annual increment in axis height for saplings for the three sites pooled was 6.42 mm.
- Immature and categories 4 and 5 growth in stem PAB and height
CAI in PAB were larger for palm trees living in ESJ. Mean CAI for PAB was 1.45 cm in PP1, 1.56 cm in PP2 and 3.61cm in ESJ. No significant differences were detected in PAB increments between PP1 and PP2. Analysis of variances indicated that there were differences for CAI in PAB between NP and ESJ (F=101.1, P<0.001). Linear regression shows no relation between CAI in PAB as independent variable and PAB as dependent.
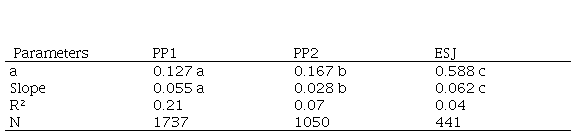
CAI in height depends on palm height and where the palm lives. There was a significant relation between increment in height and palm height. Intercepts and slops in the 3 regressions were different and grater in ESJ (Table 2).
Table 2. Parameters of the Regressions between annual increments in height as independent variable and initial height (H) as dependent variable, for Immature and categories 4 and 5. Intercepts F=100.85, P<0.001. Regression slopes F=-1047.1, P< 0.001. Tukey and Student tests: ESJ-PP2 q=18.3, p<0.001; ESJ-PP1 q=14.1, p<0.001; PP1-PP2 q=19.9, p<0.001. PP1 and PP2, permanent plots in the Iguazu National Park. ESJ, harvested area. Different letters indicate significant differences
- Categories 4 and 5: growth in PBH and PBH-height relationship
The minimum PBH measured was 10.7 cm in the park, and 13 cm in ESJ. The maximum PBH was 61 cm in the park and 54.3 cm in ESJ. In these categories, individuals developed a woody trunk with a measurable PBH when their stems reached about 2 m in height.
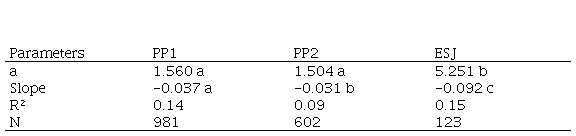
CAI in PBH was higher in ESJ palms. Regressions between CAI and PBH have negative slopes, so the wider the trunk, the lower the increment in PBH (Table 3).
Table 3. Parameters of the regression between annual stem perimeter increments and stem perimeter (PBH) for E. edulis palms in categories 4 and 5. Intercepts: F=100.85, P<0.001; slopes F=-1047.1, P<0.001), Tukey and Student test between plots (ESJ-PP2 q=-17.7 p<0.001; ESJ-PP1 q= 8.2, p<0.001; PP1-PP2 q=5.0 p<0.05). PP1 and PP2, permanent plots in the Iguazu National Park. ESJ, harvested area. Different letters indicate significant differences
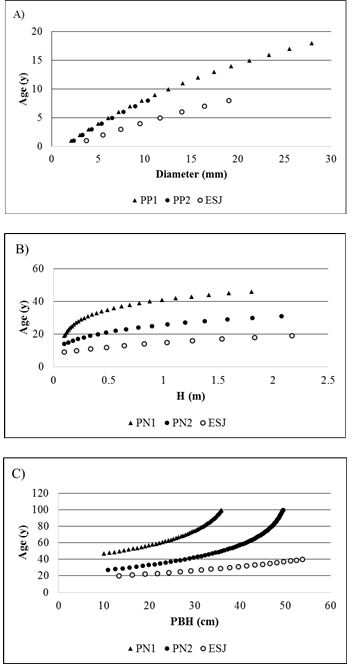
The relationship between height and PBH was a logarithmic relation. There were no differences among regressions height-PBH parameters from PP1 and PP2. Regression from ESJ was significantly different from that of the park [PBH(ESJ)=-7.18+21.08(logH(ESJ)); (R²=0.8, P(r)<0.001; P(a)<0.001, P(b)<0.001)]. National Park [PBH(NP)=-5.2+18.6(log H(NP)); (R² =0.7, P(r)<0.001; P(a)<0.001, P(b)<0.001)].
When comparing individuals of the same height, palm trees growing in the park have less stem diameter than those in the harvested area (Fig. 2).
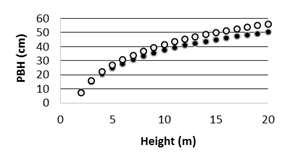
Fig. 2: Logarithmic models relating palm height and stem perimeter: in harvested area (grey dots), and protected areas (black dots).
Age estimation: Palm trees of the same size, of all categories, in harvested forests were younger than those of the same category in the park.
The larger sapling diameter was 17 mm in PN and 20 mm in ESJ. The relation between Dmax and BA is given by Dmax=20.4+3.4(lnBA); R²=0.9.
S are less in harvested areas (Table 4).
Sapling’s axis diameter in which individuals begin to develop a woody stem was the same in PP1 and PP2, so it was 26.8 mm in NP and 19.7 mm in ESJ.
In the three locations minimum immature height measured in the field was 0.1 m and the maximum was 2.0 m. The age I varies with BA, being less in harvested than in protected areas (Table 4).
The minimum PBH (PBHmin) measured were about 10.3 cm and 10.4 cm in the National Park plots, and 13.5 in ESJ. Relation between PBH minimum and BA is given by PBHmin=13.16-1.5(lnBA); R²=0.9.
The relation C=21*e(0.2AB) (R²=0.9), was used to estimate the age C of a palm that attains 10 cm in DBH under different BA. In the national park, C can double in value estimated for the harvested areas (Table 4).
Table 4. Palm trees´ age, in each category, varies in relation to category 5 density. PP1 and PP2 are permanent plots in Iguazu National Park. P0: Areas without category 5 palms. P60: Areas with 60 adult palms per ha-1. S: Age at which individuals become an immature palm. I: Age at entering in category 4. C: Age of a palm attaining a 10 cm DBH
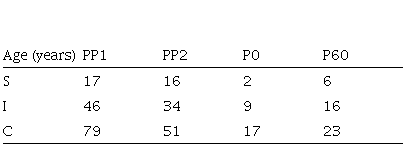
Palm trees of the same size, growing in forests with different basal areas, are different in age (Figs. 3, Table 4).
4. DISCUSSION
E. edulis individuals followed the same general palm development pattern suggested by Tomlinson (1990) and Alves et al. (2004), however their growth rates were different depending on the environmental conditions. Trees, mostly juveniles, growing in a highly shaded environment showed very low diameter growth rate and remained in the understory for a longer time increasing the risk of damage from falling debris; more light would result in faster growth that in turn would increase the chances of a tree to reach the canopy (Rozendaal et al., 2010). It seemed that this palm tree also has the ability to respond to a less crowded and more illuminated environment by increasing its size. In the harvested area, in all categories, palm trees tended to grow faster than in unmanaged mature forests.
Growth also differed under the contrasting conditions. In both areas, axis growth occurred first; height growth begun once the tree had developed a woody stem. In the harvested area, however, when saplings became immature they presented a small stem diameter, but afterwards, stem increment was higher than palm trees growing in the park. There could be different reasons for which palm trees in ESJ developed a woody stem with smaller axis diameter. In palm trees, a single stem internode is associated with each new leaf, more light radiation favour higher leaf production, which in turn increases stem development to produce new stem tissue to accommodate leaf attachment and vascular connections (Rich, 1986). Another factor could be a response to herbivore insects because in harvested areas there was a significant higher incidence of Rhyncophorus palmarum (Curculionidae) that ate saplings and immature palm heart (Gatti, 1999). Individuals that produced a woody stem earlier could have had higher probabilities to survive. Light and insect herbivore might not be the only causes for this process, there could be other factors that induce an early stem hardening.
Once the woody stem is developed, upward growth is basically influenced by the available resources. Rich (1986) recognized that palm trees that become tall at earlier stages can result in an unstable plants due to an unfavorable diameter-height relationship. Individuals living in the park tended to overbuild the stem base during the sapling stage developing a larger radicular system, which probably, allowed them to capture the limited resources in crowded environment. Developing a substantial base is necessary to have a firm stem capable of reaching the canopy slowly, or fast if gaps are present. Palm trees in ESJ after the sapling stage, had a subsequent fast increase in stem diameter which may offset the need for support to avoid falling. In fact, palms in categories 4 and 5 had a larger PAB and DBH than in the park. These results suggest that forest managers should expect a faster growth rate in the remaining individuals after the harvest. Studies in Brazil show that there is a relation between heart of palm weight and palm DBH (Reis et al., 2000). Palm trees harvested in managed areas have larger DBH and more yields per palm. However, after a palm reaches the legal diameter for cutting, DBH increment slows down. Therefore, in economic terms, is not convenient to delay harvesting once DBH reaches 10 cm. Based in this study, the legal limit of 10 cm DBH for harvesting in Argentina would be appropriate.
Traditionally peasants harvest palm trees every eight years. However, results suggest that sustainable harvest cannot be achieved in that short period of time. Chediack (2008a) suggested that palm trees harvest should be done every 13 to 20 years to allow wide population fluctuations. The amount of individuals and the conditions of harvesting must be set for each property depending on the environmental situation.
We hope that these results would increase researches interest for non-timber resources. With respect to the sustainable management of this palm tree in particular, more studies are needed, especially on harvest effects on flora and fauna diversity and frost damage, and population growth rates. Another important issue that requires attention is R. palmarum as the tree principal herbivore which can also act as a vector of a nematode that is the main plague in palm crops in Brazil and other countries.
Sustainable management of E. edulis will only be attained if illegal cuttings are stopped. Under illegal scenarios, the forests lose their economic and biological value, and owners are forced to sell their properties or transform the forest into agricultural fields. We hope that this paper contributes to understand that it takes many years of biological processes to develop an “organic” heart of palm to be served in a gourmet menu. Beyond this, there are stakeholders that believe it is possible to manage their forest in a sustainable and economically feasible manner.
5. CONCLUSIONS
1) Palm tree growth rate and form depends on environmental conditions.
2) Individuals of E. edulis respond to a less crowded and more illuminated environment, as in a harvested forests, increasing their growth rate. As saplings, they become immature at a younger age than those in the park, but with a smaller stem diameter. As immature and adults, remaining palms after harvest, have more stem increments. In protected forests, palms remain more years as saplings and tend to overbuilt theirs stem base.
3) Palms of the same size, growing in forests with different basal areas, could have different ages depending where they live. Individuals found in the logged areas reach a 10 cm stem DBH (legal limit for cutting a palm) in 23 years or less, while in protected areas they reach this size at 50 years or more.
4) The amount of individuals and time between harvests must be set for each property depending on the environmental situation.

