










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRelaciones
versão impressa ISSN 0325-2221versão On-line ISSN 1852-1479
Relaciones vol.47 no.2 Buenos Aires dez. 2022
http://dx.doi.org/10.24215/18521479e039
10.24215/18521479e039
Articulos
EXPLOTACIÓN DE AVES VOLADORAS EN LA ESTEPA DE PATAGONIA CONTINENTAL AUSTRAL: CAUQUENES (CHLOEPHAGA SP.) EN EL SITIO LAGUNA LAS VEGAS (CUENCA MEDIA DEL RÍO COYLE, ARGENTINA)
EXPLOITATION OF FLYING BIRDSIN THE STEPPE OF AUSTRAL CONTINENTAL PATAGONIA: CAUQUENES (CHLOEPHAGA SP.) IN THE LAGUNA LAS VEGAS SITE (MIDDLE BASIN OF THE COYLE RIVER, ARGENTINA)
Juan Bautista Belardi1
Luciana Stoessel2 3
1Universidad Nacional de la Patagonia Austral, Unidad Académica Río Gallegos (ICASUR), Laboratorio de Arqueología Dr. Luis A. Borrero, CONICET. E-mail: juanbautistabelardi@gmail.com
2INCUAPA, UNCPBA-CONICET, Olavarría, Buenos Aires, Argentina.
3Universidad Nacional del Centro de la Provincia de Buenos Aires (UNCPBA), Facultad de Ciencias Sociales, INCUAPA, Olavarría, Buenos Aires, Argentina. E-mail: lstoesse@soc.unicen.edu.ar
Fecha de recepción: 13 de mayo de 2022
Fecha de aceptación: 25 de agosto de 2022
RESUMEN
Se analizan los restos óseos de cauquén (Chloephaga sp.) recuperados en el sitio arqueológico a cielo abierto Laguna Las Vegas, cuenca media del río Coyle (Santa Cruz). Se evalúan los hallazgos a la luz de información arqueológica y tafonómica suprarregional, regional y local. La frecuencia de restos (NISP=23), junto con el patrón de partes esqueletarias con evidencias de consumo y las categorías etarias (cuatro individuos adultos y dos juveniles), sugieren que los cauquenes habrían sido obtenidos en proximidades del sitio durante la estación reproductiva y que se habrían transportado a este las partes de mayor rinde económico junto con las unidades anatómicas asociadas. Nuevos fechados radiocarbónicos realizados sobre un resto de cauquén y otro sobre guanaco (Lama guanicoe) sustentan la contemporaneidad aproximada en la depo-sitación de los materiales arqueológicos en el sitio alrededor de 3200 años AP.
Palabras clave: Patagonia austral - estepa - cauquenes - tafonomía - explotación -cazadores-recolectores
ABSTRACT
The results of the study carried out on cauquén -goose- (Chloephaga sp.) bone remains recovered from the open-air archaeological site of Laguna Las Vegas, middle basin of the Coyle River (Santa Cruz), are presented. We evaluate our findings in the light of supra-regional, regional and local archaeological and taphonomic information. The frequency of remains (NISP= 23), together with age patterning in skeletal parts displaying evidence for consumption (four adults and two juveniles), suggest that the cauquén would have been obtained near the site during the reproductive season, and that the parts with the highest economic yield were subsequently transported to the site together with the associated anatomical units. New radiocarbon dating ofone cauquén bone plus another on a guanaco bone (Lama guanicoe) suggest approximate contemporaneity of the archaeological materials at the site, which date to around 3200 years BP
Keywords: Southern Patagonia - steppe - cauquenes - taphonomy - subsistence - hunter-gatherers
INTRODUCCIÓN
Any one who travels through Patagonia cannot fail to be struck by the enormous quantities of upland geese (Chloephaga magellanica) which abound in the vicinity of the rivers and lagoons. At this time a great many of the birds are paired, but at a latter date in the valley of the Coyly we once made a camp round which the country in all directions was covered by thousands of these geese (Prichard 1902:76).
El comentario del periodista y viajero H. H. Prichard, realizado hace más de 100 años en el valle del río Coyle, sigue siendo absolutamente pertinente. Uno podría pensar en la importancia de estas aves como recurso para las poblaciones cazadoras-recolectoras, sin embargo, las evidencias arqueológicas de su consumo en el interior de la estepa de la Patagonia austral continental son muy escasas. Esto es así para el caso de todas las aves voladoras cuyos restos fueron interpretados como el resultado del aprovechamiento ocasional/oportunista (Humphrey et al. 1993; Cruz 2007; Mena Larraín 2009). A la luz de este marco se presenta información sobre el registro del aprovechamiento de restos de cauquenes (Chloephaga sp.) en el sitio arqueológico Laguna Las Vegas, datado en torno a 3200 años AP, en la cuenca media del río Coyle (provincia de Santa Cruz, Argentina). Es un sitio a cielo abierto ubicado sobre la margen oeste de una laguna que fue identificado a partir de la presencia de materiales óseos y líticos expuestos en superficie por erosión. Los restos se encuentran formando parte de un conjunto arqueofaunístico que se destaca tanto por la alta frecuencia de restos de guanaco (Lama guanicoe) como por su alta resolución e integridad (Stoessel et al. 2021). Debe señalarse que el guanaco ha sido la presa principal de las poblaciones cazadoras-recolectoras del interior de Patagonia desde fines del Pleistoceno (Rindel et al. 2020 y citas allí presentes).
El registro de cauquenes en el sito Laguna Las Vegas no solo es novedoso por la escasa evidencia de aves voladoras en contextos arqueológicos de Patagonia continental, sino por darse en un sitio a cielo abierto, en una alta frecuencia, con un claro perfil anatómico y tener, además, evidencias directas de procesamiento. Así, se suma información sobre la subsistencia de poblaciones cazadoras-recolectoras en la cuenca del río Coyle.
A continuación se presenta el contexto arqueológico del sitio Laguna Las Vegas en el marco regional de la arqueología de la cuenca media del río Coyle, se brindan datos ecológicos sobre Chloephaga sp., los antecedentes arqueológicos de su registro en la estepa interior de Patagonia Austral y los marcos de referencia que permiten ponderar la información recuperada en el sitio. En este sentido, se informan los resultados de observaciones tafonómicas focalizadas sobre restos de cauquén realizadas en torno a la laguna. Luego, se describen los restos recuperados en el sitio Laguna Las Vegas y se brinda un fechado taxón que permite ajustar, junto con otra nueva datación hecha sobre un resto de guanaco (Lama guanicoe), la evaluación del contexto de depositación de huesos y artefactos líticos. Por último, se evalúan y discuten los resultados del aprovechamiento de cauquenes en el marco de la arqueología de la cuenca del río Coyle y del sur de la estepa interior de Patagonia continental.
La cuenca del río Coyle está formada por numerosos tributarios cuyas nacientes se encuentran en las estribaciones orientales de la cordillera de los Andes. El río fluye por pequeños cauces de poca profundidad, lo que permite la irrigación de amplios sectores del piso de valle. Un poco más del 25% de este es ocupado por mallines con una muy buena disponibilidad de agua y amplia cobertura vegetal (alrededor del 75%) (Mazzoni y Vázquez 2004).
Laguna Las Vegas es un sitio arqueológico a cielo abierto localizado en el sector oeste de la laguna homónima, a unos 10 m de su borde (figuras 1 y 2). La laguna, formada en un bajo de origen eólico, se encuentra sobre una terraza fluvial y tiene unos 1.170 m en sentido norte-sur y 1.100 m en sentido este-oeste. Es de régimen pluvionival y tiene marcadas variaciones interanuales (Mazzoni y Vázquez 2001). Las condiciones actuales en el área se establecen hace ca. 4.000 años, cuando se instalan las comunidades vegetales de la estepa graminosa (Mancini et al. 2005). El área ecológica corresponde al matorral de mata negra (Junellia tridens), con temperaturas medias anuales entre 6,5 y 8,5 °C y lluvias entre los 150 y los 200 mm que presentan un máximo invernal (Oliva et al. 2001).
El material arqueológico se recuperó mayoritariamente de superficie y fue recolectado a lo largo de 16 m2. Dicho material proviene de un nivel estratigráfico que se encuentra entre los -12 y los -23 cm y que fue re-expuesto por la erosión del suelo. Además, se excavó una cuadrícula de un metro cuadrado en el sector de mayor frecuencia de materiales en superficie (cuadrícula metro 1 Norte) (Stoessel et al. 2021). El sitio se destaca por la alta frecuencia de individuos de guanaco (Lama guanicoe; MNI=15, NISP=333), la presencia de machacadores óseos manufacturados en restos de esta especie y un conjunto lítico compuesto principalmente por desechos de talla, raspadores y raederas confeccionados con rocas de disponibilidad local y regional. El conjunto presenta una alta integridad y resolución producto del rápido entierro de los materiales en sedimentos eólicos. Dos dataciones radiocarbónicas obtenidas sobre restos de guanaco, 3050±90 y 3210±140 años AP (tabla 1), son estadísticamente indistinguibles y muestran la contemporaneidad aproximada de la depositación de los materiales arqueológicos (Belardi et al. 2021; Stoessel et al. 2021). Los estudios sugieren que en el sitio se habrían llevado a cabo tareas de procesamiento, consumo y descarte de guanacos que habrían ingresado principalmente enteros como producto de eventos de caza atricional (Stoessel et al. 2021). A esto se suma la presencia de restos de cauquén, objeto de este trabajo.
Tabla 1. Dataciones radiocarbónicas del sitio Laguna Las Vegas. Las dataciones fueron calibradas usando el programa CALIB 8.2 (Stuiver et al. 2021) y la curva de calibración para el hemisferio Sur SHCal20 (Hogg et al. 2020) considerando dos sigmas
| Proveniencia de la muestra | Especie datada | Datación radiocarbónica (años AP) | Referencia Laboratorio | Referencias |
| Metro 1 Norte | Fémur de guanaco (Lama guanicoe) | 3210± 140 (2971-3814 cal AP) | LP-3618 | Belardi et al. 2021; Stoessel et al. 2021 |
| Metro 6 Sur | Fémur de guanaco (Lama guanicoe) | 3050 ± 90 (2938-3436 cal AP) | LP-3647 | Belardi et al. 2021; Stoessel et al. 2021 |
| Metro 1 Norte 2°nivel-23 | Húmero de guanaco (Lama guanicoe) | 3030 ± 60 (2969-3353 cal AP) | LP-3712 | Este trabajo |
| Metro 6 Norte | Húmero de cauquén (Chloephaga sp.) | 2974 ± 34 (2961-3211 cal AP) | AA-114985 | Este trabajo |
La ubicación del sitio Laguna Las Vegas difiere de la mayor parte de la información arqueológica registrada regionalmente ya que esta proviene de las terrazas fluvioglaciares que enmarcan el valle. Allí, en lagunas formadas en cuencas endorreicas -lugares reparados que concentran recursos-, se registran concentraciones de artefactos líticos producto del aprovechamiento de las rocas inmediatamente disponibles. Por fuera de las lagunas, las frecuencias artefactuales decrecen marcadamente con la excepción del registro de eventos de talla relacionados con el aprovechamiento de rodados de buena calidad.
Se propuso que el paisaje arqueológico regional sería en parte resultado de la circulación de las poblaciones cazadoras entre los valles de los ríos Santa Cruz y Gallegos, ubicados al norte y al sur del Coyle, respectivamente. Es recién en momentos de contacto donde la cuenca muestra un incremento en la residencialidad asociada a la ocupación de poblaciones Tehuelche-Aonikenk (Belardi et al. 2021).
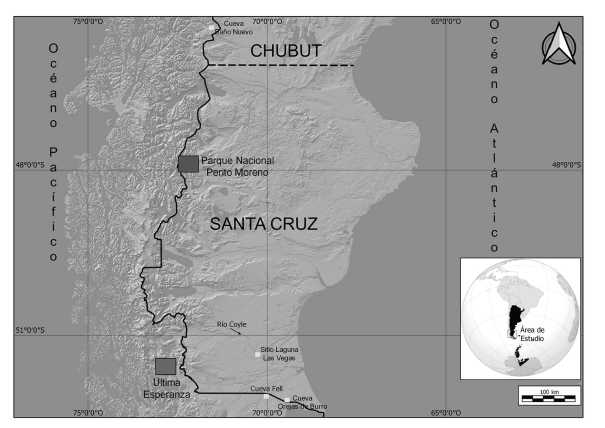
Figura 1. Localización del sitio Laguna Las Vegas y de los contextos arqueológicos de Patagonia continental austral mencionados en el trabajo

Figura 2. Sector oeste de la laguna con detalle del sitio Laguna Las Vegas y del sector excavado
EL CAUQUÉN: APUNTES ECOLÓGICOS Y ARQUEOLÓGICOS
El género Chloephaga está conformado por cuatro especies endémicas que habitan la Patagonia argentina y chilena. Tres de ellas (C. rubidiceps, C. picta, y C. poliocephala) se encuentran en el interior del territorio, mientras que la restante (C. hybrida) se restringe a la costa marina (Pedrana et al. 2011, 2018; Cossa 2019). Las especies continentales ocupan los ambientes más productivos de la estepa patagónica, principalmente las áreas de vegas o mallines ligadas a cuerpos de agua (figura 3 A). Son especies gregarias, se las suele encontrar en grupos familiares o en parejas durante la estación reproductiva (Giardina 2010). Los cauquenes presentan un dimorfismo sexual marcado y sus tamaños varían de acuerdo con la especie. El cauquén común (Chloephaga picta) es la de mayor tamaño y peso (60-65 cm; hembras: 2,47-3,05 kg, machos: 2,65-3,56 kg), seguido por el cauquén real (C. poliocephala; 50-60 cm; hembras: 1,5 kg, machos: 1,62-2,27 kg) y el cauquén colorado (C. rubidiceps; 45-52,5 cm; hembras: 1,20-1,5 kg, machos: 1,65-2,02 kg. (Cossa 2019; figura 3 B-D).
El cauquén es un ave migratoria que permanece en Patagonia durante el período reproductivo que se extiende entre septiembre y marzo. Durante este, se produce una única postura cuyo pico se desarrolla a mediados de octubre y el de nacimientos a mediados de noviembre (Cruz 2000; Pedrana et al. 2018). El tamaño de puesta más frecuente de C. picta es de seis huevos (Ibarra et al. 2010; Cossa 2019). Entre los meses de abril y mayo se traslada hacia las áreas de invernadas ubicadas al norte de Patagonia y sur de la provincia de Buenos Aires, donde se lo encuentra hasta agosto-septiembre (Pedrana et al. 2011).
Las mayores frecuencias y diversidad de restos de aves voladoras han sido registradas en el sitio Cueva Baño Nuevo-1, ubicada en la estepa del sector chileno de Patagonia centro-sur (figura 1), que tiene ocupaciones entre los 9500 y 2800 años AP (Mena Larraín 2009). Se destacan los restos de cauquenes (Chloephaga picta: N=168; Chloephaga poliocephala: N=87), varios con huellas de corte. Sobre la base de la alta frecuencia de restos de aves, se propuso que estas habrían ocupado un lugar importante en la dieta de las poblaciones humanas que habitaron este sitio y que la presencia de cuerpos de agua en su entorno posiblemente haya favorecido la presencia de este tipo de especies (Mena Larraín 2009).
En los demás contextos arqueológicos del interior de la Patagonia continental austral, el registro de cauquenes es muy escaso. En el Parque Nacional Perito Moreno (Santa Cruz, Argentina) (figura 1), un ambiente lacustre lindante con la cordillera, se han trabajado sitios en la estepa y en el ecotono con el bosque cuya cronología abarca todo el Holoceno y se ha registrado una muy baja frecuencia de restos óseos de aves voladoras (Cruz 2007, tabla 1). En la estepa, la información proviene del sitio Alero Destacamento Guardaparque (NISP=4) y del Alero Gorra de Vasco (NISP=3), mientras que en el ecotono, de las cuevas Cerro Casa de Piedra 5 (NISP=4) y Cerro Casa de Piedra 7 (NISP=63). Este último sitio es el único donde los restos de aves muestran evidencias de actividades humanas, principalmente huellas de corte y un caso de aserrado perimetral sobre una ulna de cóndor (Vultur gryphus) (Cruz 2007, figura 3) y donde se registraron restos de cauquén (NISP=10) que corresponden a las extremidades anteriores y la cintura escapular (Isabel Cruz, comunicación personal). Estas frecuencias de restos de aves no se deberían ni a problemas de muestreo ni de preservación, sino que reflejan -aparte de la posible incidencia de carnívoros-la escasa importancia de las aves voladoras en la dieta u otras actividades de subsistencia de las poblaciones cazadoras-recolectoras (Cruz 2007).

Figura 3. A. Cauquenes (C. picta) en cercanías del sitio Laguna Las Vegas; B. pareja de cauquén común (C. picta); C. pareja de cauquén real (C. poliocephala); D. pareja de cauquén colorado (C. rubidiceps)
También se han recuperado restos de cauquén en Cueva del Milodón, Alero del Diablo y Cueva Fell, Chile (Humphrey et al. 1993) (figura 1). En los dos primeros casos, ubicados en Última Esperanza, hay una muy baja frecuencia de restos de Chloephaga picta (N=3 -fragmento de cráneo y plumas-, MNI=1 y N=1 -vértebra, MNI=1, respectivamente). En la cueva del Milodón los restos corresponden a un depósito de 2500 años AP denominado conchero Nordenskjold (Luis A. Borrero, comunicación personal) y el del Alero del Diablo no tiene ni precisión cronológica ni estratigráfica. Para ninguno de los sitios se brinda información acerca del agente que aportó los restos de aves. Por su parte, en la Cueva Fell, que se encuentra en el campo volcánico Pali Aike, la cantidad de especímenes es mayor así como la diversidad de partes esqueletarias representadas (N=24, MNI=14), correspondientes a coracoides, húmeros, ulnas, carpometacarpos, sinsacro y tibiotarsos. Además, se recuperaron restos de húmero, ulna y tibiotarso que pertenecen a C. rubidiceps (N=4, MNI=3). Los restos se distribuyen a lo largo de una columna estratigráfica de más de 9030 años AP y se presume que los cazadores fueron el agente principal que ingresó los restos de aves al sitio (Humphrey et al. 1993).
En la cueva Orejas de Burro (figura 1), en Pali Aike argentino, L'Heureux (2008) identificó una alta frecuencia de restos de aves voladoras (NISP=159), que incluye tres restos de cauquén: dos radios, uno con huellas de corte, y un carpometacarpo (Lorena L'Heureux, comunicación personal). Dicha alta frecuencia se relacionaría con la cercanía del sitio al estrecho de Magallanes (17 km), principalmente a comienzos del Holoceno tardío (3500 años AP), dado que hay una importante representación de especies de aves marinas (L'Heureux 2008, tabla 3).
Las bajas frecuencias de restos de cauquenes, así como de aves voladoras en general, en los contextos arqueológicos de Patagonia austral continental quedan aún más de manifiesto cuando se las compara con la representación de restos de guanaco. Esto sugiere un comportamiento oportunista -su aprovechamiento depende de su disponibilidad- en la incorporación de las aves voladoras por parte de las poblaciones cazadores-recolectoras del interior de Patagonia continental (Cruz 2007). Otra particularidad es el predominio de restos de las extremidades anteriores y que todos los contextos reseñados corresponden a abrigos rocosos.
Las dos características del registro de cauquenes recién mencionadas también se observaron en la estepa del norte de la isla grande de Tierra del Fuego, sobre el valle del río San Martín (a unos 240 km al sur del sitio Laguna Las Vegas). Debe señalarse que dicha estepa es la continuación (interrumpida por el estrecho de Magallanes) de la estepa austral continental. En el componente A del alero Cabeza de León (post 1100 años AP) (Borrero 1979) se registraron 13 restos de cauquén que dan cuenta de al menos cuatro individuos y, si se consideran también los restos no identificados a nivel específico (NISP=30) -que podrían corresponder a Chloephaga sp. o a Anas sp.-, alcanzarían a siete (Borrero 1986). El predominio de elementos de las extremidades y el hecho de que se encuentran fracturados sugiere que no debiera descartarse un fin no económico -manufactura de cuentas- para su transporte al sitio (Borrero 1986). Unos 15 km al oeste del alero Cabeza de León se encuentra el cerro de Los Onas, Chile, que brinda reparos rocosos (Massone et al. 1993; Massone 2004). En los niveles de ocupación tempranos de la Cueva Tres Arroyos 1, ca. 10600 años AP, se registraron ocho restos de ave junto con dos epífisis de ave indeterminada y un húmero de cauquén con corte transversal; los tres interpretados como artefactos (Massone 1988, 2004). En los niveles tardíos (desde 1740 años AP hasta momentos Selk'nam) se contabilizó un MNI de 15 aves que incluye siete individuos de Chloephaga picta y dos de Chloephaga poliocephala. Además, se encontraron cuatro cuentas en huesos de ave y dos restos posibles de ser interpretados de la misma manera -uno podría ser una preforma de cuenta- (Massone 1988), lo que refuerza la sugerencia de Borrero recién referida. Asimismo, se registraron dos punzones confeccionados sobre tibiotarsos de ave, uno de ellos de cauquén en un nivel posterior a los 700 años AP (Massone 1988). El aprovechamiento de aves podría relacionarse con el complemento del guanaco durante invierno y primavera, por su mayor disponibilidad como recurso o por necesidades alimentarias relacionadas con actividades sociales significativas (Massone et al. 1993). Otros dos aleros de la localidad también tienen restos de aves. El sitio Tres Arroyos 4 presenta un NISP de 25 de los cuales 19 restos corresponden a cauquén (cráneo, húmero, radioulna, dígito mayor y menor, coracoides, fémur, tibiotarso y tarsometatarso), los que dan cuenta de tres individuos y sustentan la propuesta del traslado de los cauquenes al sitio como piezas enteras. Los restos, con excepción del nivel superior, se encuentran presentes a lo largo de toda la estratigrafía, principalmente en los niveles inferiores V y VI, considerados como pre Selk'nam (Massone et al. 1993). En el sitio Tres Arroyos 7 se registraron cinco restos de Chloephaga sp. (húmero, radio, carpometacarpo y tarsometatarso) que representan dos individuos (NISP de aves=12). Estos habrían sido consumidos en el sitio como producto de ocupaciones poco prolongadas por parte de pocos cazadores, tanto en momentos pre Selk'nam como Selk'nam -datación radiocarbónica de 100±50 años AP en el nivel III- (Massone et al. 1993). Para Tres Arroyos 1 se propusieron condiciones de preservación entre regulares y buenas, y buenas para los sitios 4 y 7 (Massone et al. 1993). En torno al faldeo norte del cerro existen más de 300 depresiones circulares que fueron interpretadas por Massone et al. (1993) como improntas de viviendas Selk'nam asignables al periodo histórico (aunque ver Ozán y Borrero [2018] donde se plantea la hipótesis alternativa de que las depresiones se deberían a procesos naturales relacionados con la deflación eólica luego de la muerte de arbustos y/o con revolcaderos de guanacos y/u ovejas). Se estudiaron dos depresiones de unos 3 m de diámetro y 30 cm de potencia arqueológica que presentaban artefactos y restos de Chloephaga sp. Para la denominada impronta 89, con fechados de 210 años AP y 280 años AP, solo se informa la presencia de restos y en la llamada impronta 88 se registró un fragmento de húmero (Massone et al. 1993). Al noreste del cerro de Los Onas se estudiaron dos sitios a cielo abierto localizados en el sistema de lagunas de las Estancias Florentina y La María, La 11 y La 12, ubicados sobre un mismo gran cuerpo de agua formado en el valle fluvioglacial sobre el que también discurre el río San Martín. En el sitio La 12 solo se encontraron restos aislados de Chloephaga sp., mientras que en el sitio La 11, interpretado como un campamento ocasional de corta duración, se identificaron restos de húmero, ulna, coracoides y tibiotarso de cauquén, que corresponderían al menos a tres individuos (presencia de tres húmeros derechos). Las presas podrían haber sido cazadas empleando trampas de lazo (Massone et al. 1993). Los restos de aves encontrados en el registro arqueológico de superficie del área de las lagunas indican el predominio de Chloephaga sp. (Massone et al. 1993).
El marco de baja frecuencia de restos de aves voladoras en el interior de Patagonia austral contrasta con el del interior de Patagonia norte. Durante el Holoceno tardío, la explotación de aves ha sido recurrente en contextos ligados a ambientes fluviales. En el valle inferior del río Chubut (unos 930 km al norte del sitio Laguna Las Vegas), en los sitios Cinco Esquinas 1 y Loma Grande 1 (ca. 1500-1200 años AP), se registró el uso casi exclusivo de recursos dulceacuícolas, entre ellos patos y/o cauquenes. El consumo de vertebrados pequeños fue interpretado como parte de un aprovechamiento oportunista en el marco de situaciones excepcionales, como las periódicas crecientes del río (Svoboda 2015). En el valle medio del río Negro se ha propuesto, sobre la base del estudio de los sitios a cielo abierto Negro Muerto y Angostura 1 (~900 años AP), que las aves habrían sido un recurso complementario y de uso recurrente durante primavera-verano (Prates y Acosta Hospitaleche 2010). Esta apreciación ha sido recientemente fortalecida y ampliada a partir de la evidencia de los sitios Pomona (Mange et al. 2020) y Colforta 1 (Mange et al. 2021), también a cielo abierto y ubicados sobre el valle medio del río Negro. El primero está datado en torno a los 1000 años AP y el segundo a los 790 años AP. Los estudios sustentan un modelo de dieta basado sobre una amplia diversidad de recursos donde las aves voladoras cumplieron un papel importante durante el Holoceno tardío (Mange et al. 2021). Una situación similar se observa en el curso inferior del río Colorado (transición pampeano-patagónica oriental). En el sitio El Tigre (ca. 900-400 años AP), ubicado en un área de médanos deflacionados en proximidades de un antiguo paleocauce, se recuperaron restos de aves con evidencias de consumo. Se propuso que la explotación de diversas especies de fauna menor formó parte del desarrollo de estrategias de intensificación y diversificación de la dieta hacia la parte final del Holoceno tardío (Alcaráz 2015).
En un reciente trabajo de síntesis, Rindel et al. (2021) proponen para el noroeste de Patagonia que las especies pequeñas, entre las que se encuentran las aves voladoras, fueron un complemento del guanaco durante el Holoceno en función de su disponibilidad y aporte nutricional y por sus menores costos de obtención y procesamiento. Estos recursos habrían facilitado el crecimiento poblacional que supera en un orden de magnitud al del sur de Patagonia (Rindel et al. 2021).
MARCOS DE REFERENCIA
La representación de partes esqueletarias de aves es el resultado de la acción conjunta de diversos agentes y procesos (Livingston 1989; Lyman 1994; Muñoz y Savanti 1998; Dirrigl Jr. 2001; Cruz 2005, 2011; Serjeantson 2009; Bovy 2012; Lefevre y Laroulandie 2014). Las observaciones tafonómicas actualistas realizadas en distintos ambientes del sur de Patagonia identificaron patrones relacionados con la representación y preservación de los conjuntos avifaunísticos de especies de tamaño mediano-grande -entre las que se encuentra el cauquén- depositados naturalmente (Cruz 2000, 2015). En general, la densidad de restos de aves voladoras en ambientes de estepa es muy baja dado que los especímenes se presentan mayoritariamente de manera aislada. A la vez, la baja frecuencia de restos está vinculada con la rápida destrucción de los especímenes en contextos de cielo abierto y en superficie, generada principalmente por la acción de carnívoros y la meteorización. A esto se suman las propiedades intrínsecas de los huesos que contribuyen a su rápida destrucción; las aves poseen huesos neumáticos, con paredes delgadas y poco resistentes a la destrucción mecánica (Livingston 1989; Cruz 2005, 2011). Asimismo, las observaciones tafonómicas registran representación diferencial de unidades anatómicas vinculada con el tipo de locomoción de las aves y con las propiedades estructurales de los huesos. En el caso las aves voladoras se registró un predominio de los elementos que conforman los miembros anteriores, seguidos por los posteriores, la cintura escapular, el esqueleto axial y la cintura pélvica. El patrón concuerda con la propuesta de diversos autores que sostienen que la anatomía funcional de las aves condiciona la morfología, estructura y densidad de los elementos afectando su resistencia a la acción de distintos procesos postdepositacionales (Livingston 1989; Cruz 2005). A la vez, puede relacionarse con la secuencia de desarticulación de las carcasas, donde los miembros posteriores lo hacen antes que los anteriores, lo que los torna más susceptibles a la acción de dichos procesos (Cruz 2011).
Los estudios actualistas evidencian que los huesos se preservarán principalmente en ambientes que favorezcan el rápido enterramiento. Es por ello que, en general, los huesos de aves en contextos estratigráficos presentan un buen estado de preservación, con escasas modificaciones y estadios bajos de meteorización (Cruz 2000, 2015). No obstante, en los sitios que están expuestos a procesos de entierro y re-exposición de materiales, la posibilidad de recuperar restos de aves disminuye y dependerá del momento de interceptación de los conjuntos.
Las actividades humanas vinculadas al aprovechamiento de las carcasas pueden generar también diferentes patrones de representación de partes esqueletarias. En el caso de las aves, si bien por su tamaño en general mediano-pequeño suelen ingresar enteras a los sitios (Lefevre y Laroulandie 2014) puede existir un aprovechamiento diferencial de unidades anatómicas de acuerdo con su potencialidad económica. En este sentido, Tivoli y Pérez (2009) desarrollaron el Índice de Utilidad de Carne Simplificado (MUI) para el cauquén común, en el que la región pectoral proporciona el mayor rendimiento, seguida por el miembro posterior (fémur y tibiotarso) y anterior (húmero). Además, para evaluar las distintas actividades vinculadas al procesamiento de las carcasas (e.g., desarticulación, descarne) es fundamental el análisis de modificaciones antrópicas como huellas de corte, fracturas y alteraciones térmicas (Laroulandie 2005; Pederg-nana y Blasco 2016). Dicho procesamiento también puede relacionarse con el empleo de otros subproductos de las aves como las plumas, lo cual puede generar patrones de modificaciones específicos (Pedergnana y Blasco 2016).
Binford (1984) observó entre los Nunamiut que cuando trozan aves del tamaño del ganso canadiense, si bien mayor que el cauquén, las alas son removidas de la pechuga como una unidad y que esta no es cortada al momento del trozamiento inicial, sino que permanece entera. Ambos aspectos tienen consecuencias arqueológicas: si las alas fueran transportadas por separado sus restos prevalecerían y el esternón permanecería entero. Esto último sería aún más esperable en presas de tamaño menor al ganso canadiense.
Por último, una excelente descripción de la caza de aves voladoras -entre otras de avutardas (cauquenes)- proviene del registro etnográfico Selk'nam, cazadores de la estepa del norte de Tierra del Fuego (Gusinde 1982). Savanti (1994) divide estos métodos de caza en el uso de armas (arco y flecha, honda y garrotes), trampas (de lazos -recordar la propuesta de Massone y coautores (1993) para el caso del sitio La 11- y de estacas con un lazo simple) y a mano. A las armas mencionadas debe sumarse el uso de bolas lanzadas de a caballo para la caza de aves pequeñas. Esto fue observado por J. B. Hatcher a fines del siglo XIX entre cazadores Aonikenk (Martinic 1995). En Patagonia continental el uso de armas arrojadizas tiene correlato material en las puntas de proyectil y las bolas, mientras que, si bien es esperable el uso de palos o ramas y de trampas, sus posibilidades de preservación son muy escasas.
Observaciones tafonómicas de cauquenes en la laguna Las Vegas
Con el fin de contar con observaciones tafonómicas directamente pertinentes en cuanto a ambiente y depositación de restos de cauquén, en abril de 2021 se realizó un relevamiento alrededor de la laguna Las Vegas. La visibilidad es excelente por lo que no hubo problemas para la identificación de restos óseos en superficie. Dos observadores recorrieron a pie todo el perímetro de la laguna considerando un ancho de relevamiento de 10 m. Las observaciones se organizaron de acuerdo con las distintas márgenes (tabla 2). La margen oeste, donde se encuentra el sitio Laguna Las Vegas, tiene 900 m de largo, una costa con sedimentos limo arcillosos, pastizales y una suave pendiente. La margen norte (900 m de longitud) tiene una playa de gravas y guijarros y una pendiente y un resalto marcados que indican la laguna. Por su parte, la margen este (1.100 m de longitud) es la más expuesta a los vientos predominantes del oeste y por eso presenta bermas muy definidas. El espacio por sobre ellas esta denudado de vegetación y es por este sector donde se realizó el relevamiento tafonómico. Por último, la margen sur tiene 800 m de longitud, una playa de guijarros y un pequeño resalto que limita la laguna y que divide la playa del espacio vegetado.
En las márgenes de la laguna se registraron restos de guanaco y aves, específicamente cau-quenes, depositados naturalmente. A los fines de este trabajo, las observaciones se focalizaron en estos últimos. Se registraron restos óseos correspondientes a 14 individuos adultos (12 cauquenes y dos indeterminados, pero de similares características). Las carcasas ubicadas en los sectores vegetados presentan un mayor grado de preservación que aquellas localizadas sobre los rodados del borde de la laguna. En todos los casos, se encuentran representados principalmente elementos correspondientes a la región pectoral y los miembros anteriores (tabla 2, figura 4). Algunas unidades anatómicas se encuentran articuladas, principalmente aquellas correspondientes a las alas, las cuales aún conservan las plumas. Los especímenes tienen escasas evidencias de meteo-rización, lo que sugiere una rápida desarticulación de los miembros posteriores. No se hicieron seguimientos sobre ellos. Las observaciones tafonómicas actualistas llevadas a cabo en la laguna Las Vegas muestran el patrón esperable para las aves voladoras de Patagonia austral (Cruz 2000, 2005) y enmarcan localmente la discusión arqueológica.
Tabla 2. Información tafonómica de la laguna Las Vegas
| Margen y superficie relevada | Individuo / especie | Elementos /Articulación / Observaciones | Sustrato | Meteorización (Muñoz y Savanti 1998) |
| Norte | 1 Cauquén | Esternón + clavículas + coracoides + fúrcula + húmeros expuestos + el resto de las dos alas con plumas | Rodados de playa de la laguna | 0-1 |
| 2 Cauquén | Esternón + clavículas + coracoides + fúrcula + húmeros expuestos + el resto de las dos alas con plumas | Rodados de playa de la laguna | 0-1 | |
| 3 Cauquén | Esternón + clavículas + fúrcula + coracoides + húmeros expuestos + el resto de las dos alas con plumas | Rodados de playa de la laguna | 0-1 | |
| 4 Cauquén | Esternón + clavículas + fúrcula + coracoides + húmeros expuestos + el resto de las dos alas con plumas | Rodados de playa de la laguna | 0-1 |
| Margen y superficie relevada | Individuo / especie | Elementos /Articulación / Observaciones | Sustrato | Meteorización (Muñoz y Savanti 1998) |
| Oeste | 5 Indeterminado | 2 húmeros / metacarpo / clavícula + coracoides + radio + ulna + metacarpo. Todos los elementos sin tejidos. Mismo individuo | Gravas sobre bermas | 1 |
| 6 Indeterminado | Radio + ulna / húmero | Gravas sobre bermas | 1 | |
| 7 Cauquén | Individuo completo con plumas. Tiene abierta la caja toráxica | Gravas sobre bermas | - | |
| 8 Cauquén | Individuo completo. Sólo las alas tienen plumas | Sector vegetado | 1-2 | |
| 9 Cauquén | Dos alas separadas con plumas / fémur | Sector vegetado | - | |
| Sur | 10 Cauquén | Esternón + clavículas + coracoides + fúrcula + húmeros expuestos + el resto del ala derecha con pocas plumas (radio y ulna expuestos). Húmero izquierdo fracturado | Sector vegetado | 1 |
| 11 Cauquén | Individuo completo con plumas | Rodados de playa de laguna | - | |
| 12 Cauquén | Húmero fracturado expuesto + resto del ala con plumas | Rodados de playa de laguna | 1 | |
| Este | 13 Cauquén | Esternón + clavículas + coracoides + fúrcula + húmeros expuestos (el derecho desarticulado) + las dos alas con plumas | Sector vegetado | 1 |
| 14 Cauquén | Individuo completo en proceso de desarticulación. Sólo las alas conservan plumas | Sector vegetado | 1 |
Referencias: + = articulación entre elementos, / = separación espacial entre elementos
LOS RESTOS DE CAUQUÉN EN EL SITIO LAGUNA LAS VEGAS
Materiales y métodos
Como se mencionó, los materiales arqueológicos del sitio fueron recuperados a partir del planteo de 16 cuadrículas de 1 m2 y de la excavación de una de ellas (cuadrícula metro 1 norte). Fueron orientadas sobre un eje oeste-este (ocho cuadrículas a cada lado del eje), de forma tal de cubrir la extensión que presentaba la mayor frecuencia de restos óseos y artefactos líticos expuestos (Stoessel et al. 2021). Los huesos de cauquén provienen de seis de las cuadrículas. En superficie se registraron 14 especímenes y dos proceden de la excavación (Stoessel et al. 2021). Asimismo, a 6, 13 y 18 m de la excavación se realizaron recolecciones de restos de cauquenes (n=7) en tres nuevas cuadrículas de 1 m2 que presentaban otros materiales arqueológicos en sectores erosionados denominados A, B y C, respectivamente. La no diferenciación de niveles estratigráficos, la representación anatómica de restos de guanaco, su meteorización y las características tecnológicas de los artefactos líticos sugieren que todos forman parte del mismo contexto de depositación que el de los demás materiales arqueológicos.

Figura 4. Carcasas de cauquenes registradas en la laguna Las Vegas; A. individuo N°5; B. individuo N°7; C. individuo N°8; D. individuo N°10; E. individuo N°11; F. individuo N°13; G. individuo N°14
En el conjunto se realizaron determinaciones y análisis cuantitativos (NISP, MNI, MAU; Grayson 1984; Lyman 1994). Las determinaciones anatómicas y de edad fueron hechas a partir de un esqueleto de referencia de un individuo adulto de cauquén que está depositado en el laboratorio de arqueología "Dr. Luis A. Borrero" y por imágenes de otro ejemplar adulto proporcionadas por la Dra. Isabel Cruz (Universidad Nacional de la Patagonia Austral). Para evaluar la integridad y resolución del conjunto se relevó la meteorización, la acción de raíces, la actividad de carnívoros y de roedores, la presencia de abrasión, pisoteo, deposiciones (e.g., óxido de manganeso, carbonato de calcio) y deterioro químico (Binford 1981; Fisher 1995; Muñoz y Savanti 1998; Behrensmeyer et al. 2003). Con el fin de ponderar los efectos de la acción antrópica se registró la presencia de huellas de corte, fracturas y alteraciones térmicas (Binford 1981; Johnson 1985; Laroulandie 2005).
Resultados
El conjunto faunístico del sitio está conformado por restos correspondientes a guanaco (n=333; 88,8%), artiodáctila (n=15; 4%), cauquén (n=23; 6,13%), ave indeterminada (n=3; 0,8%) y roedores (n=1; 0,26%; Stoessel et al. 2021). Casi en su totalidad los especímenes de cauquén corresponden a los miembros delanteros y a la región pectoral (tabla 3 y figura 5). A partir de los restos recuperados no se pudo determinar a qué especie pertenecen por lo que fueron asignados al género Chloephaga. En el conjunto se encuentran representados cuatro individuos adultos y dos juveniles, estos últimos determinados a partir del tamaño pequeño de un coracoides y de dos húmeros (figura 6). Sobre la base de la etología del cauquén la presencia de juveniles indicaría que la ocupación del sitio habría tenido lugar al menos durante los meses de primavera y verano.
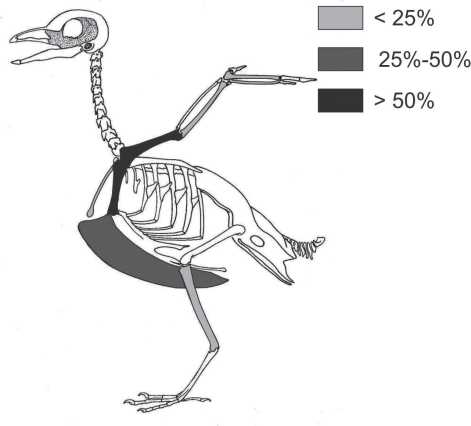
Figura 5. Representación de partes esqueletadas de Chloephaga sp.

Figura 6. Húmeros y Coracoides de Chloephaga sp. recuperados en el sitio. En cada unidad anatómica, el elemento de la izquierda corresponde a un animal adulto y el de la derecha a una cría/juvenil
Tabla 3. Partes esqueletadas correspondientes a Chloephaga sp.
| Unidad anatómica | Edad | Ubicación / cuadrícula- sector | Porción representada | Lateralidad | Huellas de corte | Meteorización (Estadio) | Marcas de raíces (% hueso afectado) | Manganeso |
| Coracoides | Adulto | Superficie/ A 16 m de la cuadrícula metro 1 Norte | Entero | derecho | - | - | - | |
| Coracoides | Adulto | Superficie/ metro 3 Norte | Entero | izquierdo | - | - | - | - |
| Coracoides | juvenil | Superficie/ A 3 m de la cuadrícula metro 5 Sur | Entero | derecho | - | - | - | - |
| Coracoides | Adulto | Superficie/ A 3 m de la cuadrícula metro 5 Sur | Entero | derecho | - | - | - | - |
| Coracoides | Adulto | Superficie/ metro 6 Norte | Entero | derecho | X | - | - | - |
| Coracoides | Adulto | Superficie/ metro 7 Norte | Ep.prox.y frag. diaf. | izquierdo | - | - | 0-25% | - |
| Coracoides | Adulto | Superficie/ metro 8 Sur | Entero | izquierdo | - | 1 | - | - |
| Húmero | Adulto | Superficie/ A 22 m de la cuadrícula metro 1 Norte | Entero | derecho | X | 2 | - | - |
| Húmero | Adulto | Superficie/ metro 6 Sur | Ep.dist.y frag. diaf. | izquierdo | - | - | - | - |
| Húmero | Adulto | Superficie/ metro 6 Sur | Entero | izquierdo | X | - | - | - |
| Húmero | Adulto | Superficie/ metro 5 Sur | Ep.prox.y frag. diaf. | izquierdo | X | - | - | X |
| Húmero | juvenil | Superficie/ A 3 m de la cuadrícula metro 5 Sur | Entero | izquierdo | - | - | 0-25% | X |
| Húmero | Adulto | Superficie/ metro 6 Norte | ep.prox.yfrag. diaf. | izquierdo | X | 2 | - | - |
| Húmero | juvenil | Superficie/ metro 7 Sur | Entero | izquierdo | - | - | - | - |
| Húmero | Adulto | Superficie/ metro 8 Sur | ep.prox. | izquierdo | - | - | 0-25% | - |
| Unidad anatómica | Edad | Ubicación / cuadrícula- sector | Porción representada | Lateralidad | Huellas de corte | Meteorización (Estadio) | Marcas de raíces (% hueso afectado) | Manganeso |
| Húmero | Adulto | Superficie/ A 16 m de la cuadrícula metro 1 Norte | Ep.dist.y frag.diaf. | - | - | - | 0-25% | - |
| Húmero | Adulto | Superficie/ A 22 m de la cuadrícula metro 1 Norte | Entero | derecho | - | - | - | - |
| Cúbito | Adulto | Superficie/ metro 1 Norte | Frag.ep.prox.y frag. diaf. | - | - | 1 | 0-25% | - |
| Carpometacarpo | Adulto | Superficie/ A 16 m de la cuadrícula metro 1 Norte | Entero | izquierdo | - | - | - | - |
| Fúrcula | Adulto | Superficie/ A 7 m de la cuadrícula metro 1 Norte | Derecha | - | - | - | - | - |
| Esternón | Adulto | Estratigrafía/ metro 1 Norte | Frag.quilla | - | - | - | 76-100% | - |
| Esternón | Adulto | Estratigrafía/ metro 1 Norte | Frag.quilla | - | - | - | 76-100% | - |
| Tibiotarso | Adulto | Superficie/ A 16 m de la cuadrícula metro 1 Norte | Frag.ep.prox.y frag. diaf. | izquierdo | - | - | - | - |
Referencias: Frag.: fragmento; ep.: epífisis; prox.: proximal; dist.: distal; diaf.: diáfisis; X: presencia
Los restos óseos presentan un buen estado de preservación, lo que refuerza las observaciones focalizadas sobre aquellos de guanaco que muestran escasas modificaciones naturales sobre sus superficies (Stoessel et al. 2021). Se registraron marcas de raíces en el 30,43% (n=7) de los especímenes, aunque la intensidad de su acción fue baja (>25% del hueso afectado). Se observaron tinciones de manganeso en dos especímenes. Solo dos restos presentan evidencias de meteori-zación en estadio 1 y dos especímenes en estadio 2 (Muñoz y Savanti 1998), lo cual sustenta la alta resolución del conjunto óseo debido a un enterramiento relativamente rápido. Esto también condice con lo observado en el conjunto de guanaco, donde los escasos restos con evidencias de meteorización en estadios 2 y 4 se corresponden con sectores expuestos de las unidades anatómicas (Stoessel et al. 2021). Con respecto a la fragmentación, doce elementos se recuperaron completos mientras que los once restantes se encuentran fragmentados. Entre estos, tres húmeros, un coracoides, un cúbito y el tibiotarso están representados por la porción proximal y diáfisis, dos húmeros por la porción distal y diáfisis, la fúrcula por la porción derecha y el esternón por dos fragmentos de quilla (tabla 3). En todos los casos se trata de fracturas postdepositacionales que, dado que la mayoría de los restos se recuperaron en superficie (tabla 3), podría deberse al pisoteo, ya sea de ovinos y/o guanacos.
Por otro lado, se registraron modificaciones en las superficies óseas que indican un origen antrópico para el conjunto. Así, cuatro húmeros y un coracoides presentan huellas de corte, que son cortas y poco profundas (figuras 7 y 8). En el caso de dos de los húmeros están localizadas en las diáfisis y dispuestas de manera transversal al eje longitudinal del hueso, mientras que en otro húmero y en un coracoides son oblicuas a dicho eje. En el cuarto húmero las huellas están sobre la epífisis proximal y se orientan de forma oblicua al eje longitudinal del hueso. La ubicación y orientación de las huellas sugiere que estarían vinculadas a actividades de descarne y desarticulación (Laroulandie 2005). No se detectaron evidencias de alteración térmica en este conjunto. Uno de los húmeros con evidencias de acción antrópica, obtenido en la cuadrícula metro 6 norte, fue datado por radiocarbono en 2974 ± 34 años (2961-3211 cal AP; AA-114985) (tabla 1). Además, se dató un húmero de guanaco con huellas de corte obtenido en el nivel inicial de depositación de materiales arqueológicos (-23 cm) en 3030±60 años (2969-3353 cal AP; LP-3712) (tabla 1).

Figura 7. A: Húmero de Chloephaga sp. con huellas de corte en la epífisis proximal; B: Húmero de Chloephaga sp. con huellas de corte en la diáfisis. Sobre este espécimen se obtuvo el fechado radiocarbónico

Figura 8. Localización de las huellas de corte en húmero y coracoides de cauquén
DISCUSIÓN Y CONCLUSIONES
Los resultados muestran que los cauquenes fueron explotados por los grupos cazadores-recolectores que ocuparon el sitio Laguna Las Vegas. La presencia de huellas de corte, así como su localización, indica el procesamiento de unidades anatómicas de alto rendimiento cárnico (Tivoli y Pérez 2009). Por su parte, las nuevas dataciones radiocarbónicas, realizadas sobre un resto de cauquén (2974 ± 34 años AP) y un espécimen de guanaco (3030 ± 60 años AP) condicen con las dos obtenidas previamente sobre huesos de guanaco, 3050 ± 90 años AP y 3210 ± 140 años AP (tabla 1). Los cuatro fechados son estadísticamente indistinguibles al 95% (t = 3.113204; x2 = 5.99; df = 2) reforzando la contemporaneidad aproximada propuesta para la depositación de los materiales arqueológicos (Stoessel et al. 2021).
El patrón de partes esqueletarias, con la presencia casi exclusiva de elementos correspondientes a la región pectoral y los miembros anteriores (tabla 3), permite pensar en dos situaciones posibles. Por un lado, que la representación de partes sea el producto de problemas de preservación diferencial y, por otro, que las presas hayan ingresado procesadas al sitio. El predominio de los elementos correspondientes a los miembros anteriores condice con lo esperable para aves voladoras, de acuerdo con las observaciones actualistas informadas en la literatura (Cruz 2000, 2005) y las aquí presentadas (tabla 2 y figura 4). Por lo tanto, el perfil de unidades anatómicas puede vincularse con sus características estructurales y anatómicas. A la vez, se destaca que este patrón se identifica tanto en sitios arqueológicos en abrigos rocosos como a cielo abierto que han tenido un rápido enterramiento, tal el caso de Laguna Las Vegas. Además, si se consideran los elementos de manera individual, el coracoides y el húmero son los que presentan la mayor frecuencia (siete y diez, respectivamente). Estos poseen una relativa alta densidad mineral ósea por lo que su probabilidad de supervivencia es mayor (Muñoz y Savanti 1998; Dirrigl Jr. 2001; Serjeantson 2009). Sin embargo, se recuperaron también especímenes que tienen baja densidad mineral como es el caso de los fragmentos de quilla y no se han registrado huellas de carnívoros, que han sido mencionados como agentes que pueden transportar restos de aves a los sitios arqueológicos así como también generar la rápida destrucción de los especímenes (Cruz 2011). Sumado a esto, el análisis tafonómico muestra que los restos poseen un buen estado de preservación asociado con su relativo rápido enterramiento. Las mismas condiciones han sido observadas en el caso de los restos de guanaco, lo que avala la alta integridad y resolución propuesta para el sitio (Stoessel et al. 2021). Los estudios actualistas mostraron que la probabilidad de supervivencia de unidades anatómicas de aves en contextos a cielo abierto de la estepa patagónica es muy baja; es en los abrigos rocosos donde se dan condiciones que favorecen la preservación (Cruz 2011). No obstante, Laguna Las Vegas evidencia que en los sitios a cielo abierto de la estepa localizados en ambientes lagunares con una pronta cobertura sedimentaria la posibilidad de recuperar restos de aves es alta, tal como es el caso de los sitios ubicados en el norte de Tierra del Fuego y en los valles de los ríos Chubut, Negro y Colorado, en Patagonia norte. No obstante, todos estos sitios tienen o permiten inferir cronologías mucho más tardías, lo que destaca las buenas condiciones de preservación de Laguna Las Vegas. Otro aspecto interesante que surge de la comparación suprarregional es que las presas de porte menor tuvieron un rol complementario al aprovechamiento del guanaco en el norte (Prates y Acosta Hospitaleche 2010; Alcaráz 2015; Svoboda 2015; Mange et al. 2020, 2021) y Noroeste de Patagonia; en este último caso con un amplio marco temporal (Rindel et al. 2021).
Entonces, sobre la base de la discusión precedente, se propone que el patrón de partes esque-letarias de cauquenes de Laguna Las Vegas es el producto principal de la selección y transporte por parte de las poblaciones cazadoras-recolectoras de presas que se habrían obtenido en las inmediaciones del sitio. Si se considera su potencialidad económica, la región pectoral es la que posee el valor del MUI más elevado, siendo el esternón el de mayor rendimiento, mientras que el coracoides tiene valores medios (Tivoli y Pérez 2009). En consecuencia, la presencia de estos elementos podría responder a la selección de aquellas unidades de mayor importancia económica. La alta frecuencia de coracoides y húmeros sugiere que ambos elementos habrían conformado la parte principal de un segmento anatómico seleccionado, el ala superior, ya que casi no hay elementos del ala inferior, de marcado menor rinde económico. Esto es en parte coincidente con las observaciones realizadas por Binford (1984) sobre el trozamiento de aves del tamaño del ganso canadiense, a la que se le suma en este caso patagónico el descarte del extremo inferior del ala. En cuanto a la conservación del esternón entero, es difícil precisarlo dada su fragilidad. No obstante, la presencia de las quillas con restos de ambos lados del esternón, se ajustaría a la expectativa de su conservación como una pieza entera.
La propuesta de obtención de los cauquenes en cercanías del sitio estriba en que el entorno de pastizales de la laguna Las Vegas reúne las características de las áreas de nidificación. Las buenas condiciones de preservación del sitio permiten postular que los cauquenes fueron obtenidos, luego descartadas las partes de menor rinde económico y aquellas de mayor rendimiento transportadas al sitio, así como las unidades anatómicas asociadas (miembro anterior superior). En este caso, también podría haber un aprovechamiento de las plumas -aunque la ausencia de los elementos distales de las alas indica que no habría sido un objetivo del procesamiento (Bovy 2012)- y, por supuesto, de sus huevos, si se tiene en cuenta que los cauquenes habrían sido obtenidos durante la estación de reproducción, si bien no hay evidencia arqueológica de ello.
En el sitio no se encontraron artefactos sobre huesos de ave y ninguno de los tipos de artefactos líticos y óseos recuperados difieren de los observados en el contexto regional, son los mismos que forman parte del "ruido de fondo tecnológico" de la cuenca media del río Coyle (Belardi et al. 2021; Stoessel et al. 2021). Esto indica que no hay artefactos que puedan relacionarse específicamente con el procuramiento de los cauquenes y refuerza la idea, anclada en la etnografía, de la posibilidad del empleo de garrotes, trampas, la caza a mano o, además, del aprovechamiento de carcasas de individuos muertos naturalmente, tal como lo muestra la información tafonómica. Esto da lugar a que la obtención de cauquenes -y la recolección de huevos- podría haber sido una actividad realizada por miembros de grupos cazadores de diferentes edades.
El registro de cauquenes en el sitio Laguna Las Vegas constituye un caso novedoso en el marco de la baja frecuencia de restos de esta especie en contextos arqueológicos del sector continental de estepa interior de Patagonia austral y, además, por ser en un sitio a cielo abierto. La evidencia sostiene que la baja frecuencia de restos de aves voladoras estaría vinculada con un aprovechamiento oportunista. No obstante, en contextos con condiciones ecológicas favorables, es probable que la incorporación de las aves voladoras a la subsistencia de las poblaciones ca-zadoras-recolectoras haya sido mayor, tal como en el caso del sitio Baño Nuevo (Mena Larraín 2009), en los distintos sitios del norte de Tierra del Fuego (Massone et al. 1993) y en los de Patagonia norte (Prates y Acosta Hospitaleche 2010; Alcaráz 2015; Svoboda 2015; Mange et al. 2020, 2021). En consecuencia, en presencia de entornos lagunares/mallines vinculados con las áreas de reproducción, la explotación de cauquenes en el interior de Patagonia continental podría haber sido un complemento usual del aprovechamiento del guanaco desde la primavera hasta comienzos del otoño.
AGRADECIMIENTOS
A la Flia. Arbilla, de Ea. Rubén Aike, por permitirnos trabajar en su propiedad. Alejandro Súnico nos informó de la concentración de arqueofauna y Flavia Carballo Marina y Pablo Binaghi nos ayudaron en los relevamientos. Isabel Cruz y Lorena L'Heureux muy gentilmente nos facilitaron datos inéditos de sus investigaciones. A Flavia Carballo Marina, Ana Paula Alcaráz, Luis A. Borrero, Matías Medina y Gustavo Martínez por la valiosa discusión de distintos aspectos del trabajo. A Carlos Albrieu por su asesoramiento sobre aves y a William Taylor por la revisión del abstract. A los evaluadores por sus observaciones y sugerencias. A la Universidad Nacional de la Patagonia Austral, Unidad Académica Río Gallegos, por posibilitar el desarrollo de la investigación.
BIBLIOGRAFÍA
Alcaráz, A. P. (2015). Aprovechamiento antrópico de fauna menor en el curso inferior del río Colorado (transición pampeano-patagónica oriental): el sitio El Tigre como caso de estudio. Intersecciones en Antropología 16: 115-129.
Behrensmeyer, A. K., Stayton, C. T. y Chapman, R. E. (2003). Taphonomy and ecology of modern avifaunal remains from Amboseli Park, Kenya. Paleobiology 29: 52-70.
Belardi, J. B., Carballo Marina, F. y Sáenz, J. L. (2021). Hunter-gatherers in a broad landscape: off-site regional archaeology in the Coyle River basin, Southern Patagonia (Argentina). En M. Bonomo y S. Archila (eds.), South American Contributions to World Archaeology: 141-157. One World Archaeology series, Springer-Nature, Cham.
Binford, L. R. (1981). Bones: Ancient Men and Modern Myths. Academic Press, New York.
Binford, L. R. (1984). Butchering, Sharing, and the Archaeological Record. Journal of Anthropological Archaeology 3: 235-257.
Borrero, L. A. (1979). Excavaciones en el alero "Cabeza de León". Isla grande de Tierra del Fuego. Relaciones de la Sociedad Argentina de Antropología XII: 255-271.
Borrero, L. A. (1986). La economía prehistórica de los habitantes del norte de la Isla Grande de Tierra del Fuego. Tesis Doctoral inédita, Facultad de Filosofía y Letras, Universidad de Buenos Aires.
Bovy, K. M. (2012). Why so many wings? A re-examination of avian skeletal part representation in the south-central Northwest coast, USA. Journal of Archaeological Science 39 (7): 2049-2059.
Cossa, N. (2019). Biología de la conservación de tres especies de cauquén (Chloephaga spp.). Tesis Doctoral inédita, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires.
Cruz, I. (2000). Líneas tafonómicas y ecológicas para evaluar la explotación prehistórica de aves acuáticas en la zona cordillerana (provincia de Santa Cruz, Argentina). En Universidad Nacional de la Patagonia Austral (ed.), Desde el País de los Gigantes. Perspectivas arqueológicas en Patagonia I: 203-218. Río Gallegos.
Cruz, I. (2005). La representación de partes esqueletarias de aves. Patrones naturales e interpretación arqueológica. Archaeofauna 14: 69-81.
Cruz, I. (2007). La explotación de las aves en la zona cordillerana del sur de Patagonia (Santa Cruz, Argentina). Revista Werken 11: 107-126.
Cruz, I. (2011). Tafonomía de huesos de aves. Estado de la cuestión y perspectivas desde el sur del Neotrópico. Antípoda. Revista de Antropología y Arqueología 13: 147-174.
Cruz, I. (2015). Las investigaciones sobre preservación de huesos de aves y mamíferos grandes en Patagonia (Argentina). Archaeofauna 24: 209-224.
Dirrigl Jr., F. J. (2001). Bone Mineral Density of Wild Turkey (Meleagris gallopavo). Skeletal Elements and its Effect on Differential Survivorship. Journal of Archaeological Science 28: 817-832.
Fisher Jr., J. W. (1995). Bone Surface Modifications in Zooarchaeology. Journal of Archaeological Method and Theory 2 (1): 7-68.
Giardina, M. A. (2010). El aprovechamiento de la avifauna entre las sociedades cazadoras-recolectoras del sur de Mendoza: un enfoque arqueozoológico. Tesis Doctoral inédita, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata.
Grayson, D. K. (1984). Quantitative Zooarchaeology: Topics in the Analysis of Archaeological Faunas. Orlando, Academic Press.
Gusinde, M. (1982). Los indios de Tierra del Fuego. Tomo I: Los Selk'nam. CAEA-CONICET, Buenos Aires.
Hogg, A. G., Heaton, T. J. Hua, Q. Palmer, J. G. Turney, C. S. M. Southon, J., Bayliss, A., Blackwell, P., Boswijk, G., Bronk Ramsey, C., Pearson, C. Petchey, F., Reimer, P., Reimer, R. y Wacker, L. (2020). SHCal20 Southern Hemisphere calibration, 0-55.000 years cal BP. Radiocarbon 62 (4): 759-778.
Humphrey, P. S., Péfaur, J. E. y Rasmussen, P. C. (1993). Avifauna of three Holocene cave deposits in Southern Chile. Occasional Papers of the Museum of Natural History, The University of Kansas Lawrence 154: 1-37.
Ibarra, J. T., Schüttle, E., McGehee, S. y Rozzi, R. (2010). Tamaño de puesta, sitios de nidificación y éxito reproductivo del caiquén (Chloephaga picta Gmelin, 1789) en la reserva de biosfera Cabo de Hornos, Chile. Anales Instituto Patagonia 38 (1): 73-82.
Johnson, E. (1985). Current developments in bone technology. En M. B. Schiffer (ed.), Advances in Archaeological Method and Theory 8: 157-235. Academic Press, New York.
Laroulandie, V. (2005). Bird exploitation pattern: the case of Ptarmigan Lagopus sp. in the Upper Magdalenian site of La Vache (Ariege, France). En G. Grupe y J. Peters (eds.), Feathers, Grit and Symbolism. Birdsand Humans in the Ancient Old and New Worlds: 165-178. Proceedings of the 5th Meeting of the ICAZ Bird Working Group, Munich.
Lefevre, C. y Laroulandie, V. (2014). Avian Skeletal Part Representation: A Case Study from Offing 2, A hunter-Gatherer-Fisher Site in the Strait of Magellean (Chile). International Journal of Osteoarchaeology 24: 256-264.
L'Heureux, L. (2008). La arqueofauna del campo volcánico Pali Aike. El sitio Orejas de Burro 1, Santa Cruz, Argentina. Magallania 36 (1): 65-78.
Livingston, S. D. (1989). The taphonomic interpretation of avian skeletal part frequencies. Journal of Archaeological Science 16: 537-547.
Lyman, L. R. (1994). Vertebrate Taphonomy. Cambridge University Press, Cambridge.
Mancini, M. V., Paez, M., Prieto, A., Stutz, S., Tonello, M. y Vilanova, I. (2005). Mid-Holocene climatic variability reconstruction from pollen records (32°-52° S, Argentina). Quaternary International 132: 47-59.
Mange, E., Di Lorenzo, M., González Venanzi, L. y Anthonioz Blanc, I. (2021). Consumo de aves en el valle medio del río Negro durante el Holoceno tardío final: los restos del sitio Colforta 1 (provincia de Río Negro, Argentina). Intersecciones en Antropología 22 (2): 169-181.
Mange, E., Fernández, F. y Di Lorenzo, M. (2020). Análisis del conjunto faunístico el sitio arqueológico Pomona (provincia de Río Negro, Argentina). Comechingonia. Revista de Arqueología 24 (3): 255-279.
Martinic, M. (1995). Los Aonikenk. Ediciones de la Universidad de Magallanes, Punta Arenas, Chile.
Massone, M. (1988). Artefactos óseos del sitio arqueológico Tres Arroyos (Tierra del Fuego). Anales del Instituto de la Patagonia. Serie Ciencias Sociales 18: 107-112.
Massone, M. (2004). Los cazadores después del hielo. Colección de Antropología. Centro de Investigaciones Diego Barros Arana, Santiago, Chile.
Massone, M., Jackson, D. y Prieto, A. (1993). Perspectiva arqueológica de los Selk'nam. Colección de Antropología. Centro de Investigaciones Diego Barros Arana, Santiago, Chile.
Mazzoni, E. y Vázquez, M. (2001). Evaluación de pastizales húmedos para un aprovechamiento sustentable en la cuenca del río Gallegos (Provincia de Santa Cruz, Argentina). VIII Encuentro Latinoamericano de Geógrafos: 175-182.
Mazzoni, E. y Vázquez, M. (2004). Ecosistemas de mallines y paisajes de la Patagonia Austral (Provincia de Santa Cruz). Ediciones Instituto Nacional de Tecnología Agropecuaria, Buenos Aires.
Mena Larraín, F. (2009). Aves en Cueva Baño Nuevo. En P. López M., I. Cartajena F., C. García P. y F. Mena Larraín (eds.), Zooarqueología y Tafonomía en el confín del mundo: 59-71. Monografías Arqueológicas N° 1, Universidad Internacional Sek-Chile.
Muñoz, S. A. y Savanti, F. (1998). Observaciones tafonómicas sobre restos avifaunísticos de la costa Noreste de Tierra del Fuego. Actas y Memorias del XI Congreso Nacional de Arqueología Argentina (8a parte). Revista del Museo de Historia Natural de San Rafael XX (1/4): 107-121.
Oliva, G., González, L., Rial, P. y Livraghi, E. (2001). El ambiente en la Patagonia austral. En P. Borrelli y G. Oliva (eds.), Ganadería ovina sustentable en la Patagonia austral. Tecnología de manejo extensivo: 19-82. Ediciones. Instituto Nacional de Tecnología Agropecuaria, Río Gallegos.
Ozán, I. y Borrero, L. A. (2018). Sobre el origen de las depresiones circulares del talud de Tres Arroyos, Tierra del Fuego. Magallania 46 (2): 167-185.
Pedergnana, A. y Blasco, R. (2016). Characterising the exploitation of avian resources: An experimental combination of lithic use-wear, residue and taphonomic analyses. Quaternary International 421: 255-269.
Pedrana, J., Bustamante, J., Rodríguez, A. y Travaini, A. (2011). Primary productivity and anthropogenic disturbance as determinants of upland goose Chloephaga picta distribution in southern Patagonia. The International Journal of Avian Science 153: 517-530.
Pedrana, J., Pütz, K., Bernad, L. Seco Pon, J. P., Gorosabel, A., Muñoz, S. D., Isacch, J. P. Matus, R., Blanck, O., Lüthi, B., Lunardelli, M. y Rojas, P. (2018). Migration routes and stopover sites of upland geese Chloephaga picta in South America. Avian Biology Research 11 (2): 89-99.
Prates, L. y Acosta Hospitaleche, C. (2010). Las aves de sitios arqueológicos del Holoceno tardío de Norpatagonia, Argentina. Los sitios Negro Muerto y Angostura 1 (Río Negro). Archaeofauna 19: 7-18.
Prichard, H. H. (1902). Through the Heart of Patagonia. D. Appleton and Company, New York.
Rindel, D. D., Moscardi, B. F. y Perez, S. I. (2020). The distribution of guanaco (Lama guanicoe) in Patagonia during Late Pleistocene-Holocene and its importance for prehistoric human diet. The Holocene. https://doi. org/10.1177/0959683620981689.
Rindel, D. D., Gordón, F., Moscardi, B. y Perez, S. I. (2021). The Role of Small Prey in Human Populations of Northwest Patagonia and its Implications. En J. B. Belardi, D. L. Bozzuto, P. M. Fernández, E. A. Moreno y G. A. Neme (eds.), Ancient Hunting Strategies in Southern South America: 175-207. The Latin American Studies Book Series, Springer.
Savanti, F. (1994). Las aves en la dieta de los cazadores-recolectores terrestres de la costa fueguina. Temas de Arqueología. Consejo Nacional de Investigaciones Científicas y Técnicas, Programa de Estudios Prehistóricos, Buenos Aires.
Serjeantson, D. (2009). Birds. CambridgeManuals inArchaeology. Cambridge University Press, Cambridge.
Stoessel, L., Belardi, J. B. y Carballo Marina, F. (2021). Sitio Laguna Las Vegas: procesamiento y consumo de guanacos (Lama guanicoe) en la cuenca media del río Coyle (provincia de Santa Cruz). Relaciones 46 (1): 339-361.
Stuiver, M., Reimer, P. J. y Reimer, R. W. (2021). CALIB 8.2 [www program] at http://calib.org
Svoboda, A. (2015). Los vertebrados pequeños en la subsistencia de cazadores-recolectores: una evaluación zooarqueológica comparativa para Patagonia central. Tesis Doctoral inédita, Facultad de Filosofía y Letras, Universidad de Bueno Aires.
Tivoli, A. y Pérez, A. (2009). Rendimiento económico del cauquén común (Chloephaga picta, Familia: Anatidae). En M. Salemme, F. Santiago, M. Álvarez, E. Piana, M. Vázquez y M. E. Mansur (eds.), Arqueología de la Patagonia: Una Mirada desde el confín del mundo 2: 853-864. Editorial Utopías, Ushuaia.

