











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Santiago del Estero es la provincia con mayor desarrollo de la ganadería en el noroeste de la Argentina, con aproxima damente el 50% del stock ganadero re gional ( Moscuzza et al., 2003). Las pastu ras, tanto gramíneas como leguminosas, constituyen la fuente principal en la die ta del ganado. Actualmente se dispone de un espectro relativamente amplio de forrajeras adaptadas a las diferentes con diciones edafo-climáticas del Chaco se miárido, sin embargo, la implantación de leguminosas incrementa la materia orgá nica de los sistemas, incorpora nitrógeno al suelo y aumenta la producción de carne por hectárea ( Lamela et al., 2009).
La implantación de leguminosas exóti cas puede presentar problemas de adap tación a las condiciones semiáridas por lo que es recomendable la utilización de especies nativas ya adaptadas a estos am bientes ( Pang et al ., 2011 ). Numerosos estudios están enfocados a introducir le guminosas nativas como forrajeras a fin de conservar y resguardar el germoplas ma nativo, mejorar la cobertura vegetal, la calidad y cantidad de forraje para el ganado y utilizar los recursos locales ( Basconsuelo et al., 2013).
En la región chaqueña semiárida, en pastizales nativos, sistemas silvopastori les y potreros donde se han implantado forrajeras megatérmicas, es frecuente encontrar leguminosas nativas consumi das por el ganado. Entre ellas, Coursetia hassleri, leguminosa perenne, semile ñosa de porte erguido y especialmente frecuente en los lugares sombreados, es considerada con potencial forrajero para la región ( Burkart, 1943). Es perenne de crecimiento primavero-estival, indeter minado, por lo que durante su ciclo pro ductivo se observa la presencia simultá nea de flores y frutos en diversos estadios de desarrollo. Durante el otoño-invierno entra en reposo, perdiendo todas sus ho jas y los órganos reproductivos. En agos to, se produce el rebrote, reanudando su crecimiento, con una longitud de ciclo de 210 días para las condiciones climá ticas de Santiago del Estero ( Ruiz, 2017).
El valor nutricional de una especie forrajera se determina comúnmente en términos de proteína bruta (PB), materia seca digestible, contenido de fibra deter gente neutra (FDN), fibra detergente áci da (FDA) y energía metabolizable ( Ar zani et al., 2003). En algunas gramíneas forrajeras el valor nutritivo va disminu yendo con la madurez de la planta. En es tas especies, la concentración de la pared celular es alta y su estructura variable, siendo ambas afectadas por el genotipo, la madurez y el ambiente ( Barrio et al., 2014). En Panicum maximum y Bothrio chloa saccharoides la calidad nutritiva aumentó al incrementar el sombreo, con menor contenido de FDN y FDA. En Panicum maximun, el rendimiento fue mayor al aumentar el nivel de exposición al sol ( Obispo et al ., 2008 ; Piñeros et al ., 2011 ). En tal sentido, las condiciones lu mínicas podrían modificar la calidad de las pasturas, factor a considerar en siste mas sombreados como los silvopastori les, sin embargo, todavía quedan interro gantes por profundizar.
Además del valor nutritivo, es necesa rio considerar la presencia de metabo litos secundarios en las plantas que ge neralmente se sintetizan como respuesta de defensa a la herbivoría y que varían según el estado fenológico y el ambiente, presentando en algunos casos un efecto antinutricional en herbívoros ( Salami et al., 2019). Entre los metabolitos se cundarios se incluyen los compuestos fenólicos, muy variables en estructura y bioactividad: ácidos fenólicos, lignanos, taninos (hidrolizables y condensados) y flavonoides ( Isaza Martinez, 2007). Ni veles de consumo de taninos condensa dos superiores a 40-50 g kg -1 de forraje, pueden reducir la digestibilidad de las proteínas y de la materia seca, afectando negativamente el consumo voluntario de alimento ( McMahon et al ., 2000 ). En dosis adecuadas, pueden mejorar el pro ceso digestivo y la nutrición al formar complejos con proteínas que evitan la de gradación de las mismas por microorga nismos ruminales, aumentando el flujo y absorción de nitrógeno no amoniacal en el intestino delgado. Asimismo, pueden evitar el timpanismo producido en ru miantes al ingerir grandes cantidades de leguminosas ( McMahon et al ., 2000 ) y reducir el parasitismo ( Torres-Acosta et al ., 2008 ), constituyendo una alternativa prometedora para reemplazar antihelmínticos sintéticos ( Quijada et al., 2015).
Aunque se han realizado algunos estu dios exploratorios en C. hassleri ( Barrio et al., 2015; Fuentes et al., 2016), no se han evaluado las modificaciones en cali dad forrajera y contenido de compuestos fenólicos a medida que transcurre su ci clo productivo.
Con la finalidad de utilizar a Coursetia hassleri como una alternativa forrajera, el objetivo de este trabajo fue evaluar el efecto de las condiciones lumínicas y el momento del ciclo productivo de la planta sobre: a) la producción de mate ria seca y calidad nutritiva de las plan tas y b) el contenido y distribución de compuestos fenólicos en hojas, tallos y frutos de C. hassleri.
Material y Método
El ensayo se llevó a cabo en el campo experimental FAyA-UNSE, localidad El Zanjón (Santiago del Estero), coorde nadas 27°52´21” S y 64°14´33” O y 188 m s.n.m., durante el ciclo productivo 2016/2017. Se utilizaron 6 parcelas de 3 x 4 m de superficie, implantadas en 2015 con C. hassleri, en un marco de planta ción de 0,30 x 0,30 m y regadas sema nalmente. Se evaluaron dos condiciones lumínicas: sol (Csol), parcelas bajo cielo abierto, y sombra (Csom), parcelas bajo media sombra de 50% de intercepción de la luz solar. En cada condición lu mínica se utilizaron tres repeticiones (3 parcelas). El efecto del momento del ciclo productivo se evaluó en diciembre (12/12/2016) cuando las plantas alcanza ron 40 a 50 cm de altura (considerando que ese tamaño representa una acumula ción de material suficiente como para ser ofrecido como alimento a los animales) y al final del ciclo, en febrero (16/2/2017). Las plantas cortadas en diciembre fue ron marcadas para no ser cortadas nue vamente en febrero, a fin de analizar la evolución de la calidad nutritiva y el contenido de compuestos fenólicos con el avance del ciclo productivo. Las mues tras se tomaron utilizando un cuadrante de 1 x 1 m, arrojado al azar en cada par cela y cortando el material vegetal a 10 cm del suelo. El material cortado fue pe sado (peso fresco) y llevado a estufa a 55 °C hasta peso constante a fin de estimar la producción de materia verde (MV) y seca (MS). Las muestras secadas fueron molidas a un tamaño de partícula de 1 mm (molino Willey modelo TS3375E15, Thomas Scientific, Swedesboro, New Jer sey, USA) y almacenadas a 25 °C hasta su análisis. Se determinaron: proteína bruta (PB) por Kjeldahl ( AOAC, 1984), fibra detergente ácido (FDA) y fibra detergen te neutro (FDN) mediante Ankom 200 Fiber Analyzer ( Komarek, 1993) y mate ria seca (%MS) de laboratorio a 105 °C.
La cuantificación de compuestos fenó licos se realizó sobre una alícuota de 100 g del material cosechado, que fue sepa rada en hojas, tallos y frutos; cada frac ción secada en estufa a 55 °C, molida y almacenada a -18 °C dentro de bolsas de polietileno hasta el momento del análi sis. Para la preparación del extracto, so bre 0,1 g de material vegetal molido se realizó una extracción con una mezcla de acetona-agua (70:30 v/v) por una hora con períodos de sonicación. Se centrifu gó 10 min a 10.000 rpm y se transfirió el sobrenadante llevando a volumen final de 10 ml. El extracto preparado se utili zó para cuantificar el contenido total de compuestos fenólicos (FT), taninos con densados extraíbles (TC), taninos totales (TT) y flavonoides. La determinación de FT se realizó por espectrofotome tría según el método de Folin-Ciocalteu ( Singleton & Rossi, 1965) y se expresó como equivalentes de Ácido Gálico (g AGa/100 g MS). Los TC fueron deter minados por espectrofotometría según el método Butanol-HCl y expresados como equivalentes de cianidina (g Cia nidina/100 g MS) ( Porter et al ., 1985 ). El contenido de TT se determinó por diferencia entre FT antes y después de la extracción de los taninos por adsorción en polivinilpolipirrolidona (PVPP) que forma un precipitado insoluble. Se midió el contenido fenólico al sobrenadante mediante el método de Folin-Ciocalteu y se expresó como equivalente de ácido gálico (mg AGa/g MS). Los flavonoides se cuantificaron por espectrofotometría, utilizando como reactivo el cloruro de aluminio (AlCl 3) ( Marcucci et al ., 1998 ), expresando los resultados como equiva lentes de quercetina (mg Q/ g MS).
Análisis estadístico
Se utilizó un diseño aleatorizado con 4 tratamientos para la determinación de producción de materia verde (MV), seca (MS) y calidad nutritiva en un análisis factorial 2 x 2: dos momentos del ciclo productivo y dos condiciones de luz. Para los análisis de compuestos fenólicos se utilizó una estructura factorial 2 x 2 x 3: dos momentos del ciclo productivo, dos condiciones de luz y tres órganos (hoja, tallo, fruto). Los datos obtenidos se anali zaron mediante MLGM con prueba DLS de Fisher al 0,05 de significancia para las diferencias entre medias, utilizando el software InfoStat ( Di Rienzo et al., 2018).
Resultados
No hubo interacción significativa en tre los factores luz y momento del ciclo ( Tabla 1).
Tabla 1: Producción de materia verde (MV; kg/ha), porcentaje (MS; %), y producción de mate ria seca (MS, kg/ha) según la condición de luz y el momento del período de crecimiento. Table 1: Fresh matter production (FM, kg/ha), percentage (DM, %) and dry matter production (DM, kg/ha) according to the light conditions and the moment of the growth period
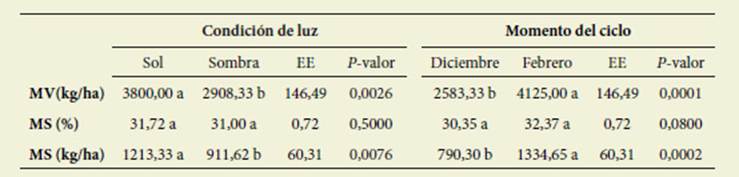
Letras diferentes indican diferencias significativas (valor p < 0,05)
Different letters indicate significant statistical differences (p value <0,05)
Con respecto al contenido de MS (%), ni la luz ni el momento del ci clo produjeron diferencias significativas (p> 0,05). Sin embargo, la producción tanto de MV como de MS fue significa tivamente mayor en luz y en febrero, al avanzar el ciclo productivo.
Para la proporción de órganos en las plantas, no se observó interacción sig nificativa entre los factores estudiados. La proporción de hojas, tallos y frutos no fue modificada por la luz, pero sí por el momento del ciclo ( Figura 1).
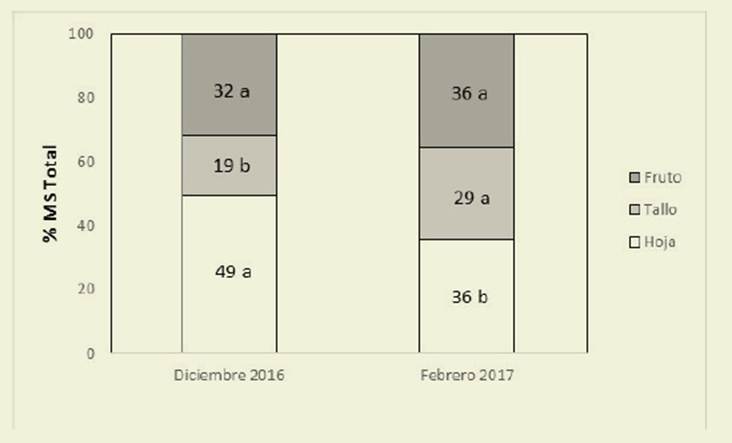
Figura 1: Proporción (porcentaje de MS total) de hojas, tallos y frutos de C. hassleri en dos momentos del ciclo del cultivo Figure 1: Proportion (percentage of total DW) of leaves, stems and fruits of C. hassleri in two moments of the plants cycle
En di ciembre la proporción de hojas fue ma yor y la de tallos menor que para febrero mientras que la de frutos no fue modifi cada a lo largo del ciclo.
No hubo interacción significativa (va lor p> 0,05) entre condición de luz y momento del ciclo para las variables de calidad nutritiva evaluadas. Ninguna de ellas fue modificada por las condiciones lumínicas, aunque sí hubo diferencias significativas según el momento del ci clo productivo ( Tabla 2).
Tabla 2: Calidad nutritiva promedio de Coursetia hassleri bajo dos condiciones de luz y dos momentos del período de crecimiento Table 2: Mean nutritional quality of Coursetia hassleri under two light conditions and two moments of the growth period
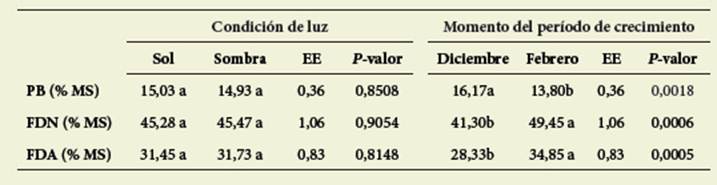
Letras diferentes indican diferencias significativas (p< 0,05).
Different letters indicate significant differences (p< 0,05).
El conteni do de PB fue significativamente mayor (valor p=0,0018) en diciembre ( Tabla 2) mientras que los valores de FDN y FDA fueron significativamente mayores (p=0,0006 y p=0,0005) en febrero.
El contenido de FT no fue afectado por el momento del ciclo y las condiciones de luz (p = 0,4741 y 0,3151 respectivamen te), pero presentó variaciones significa tivas en hojas, tallos y frutos ( Figura 2).
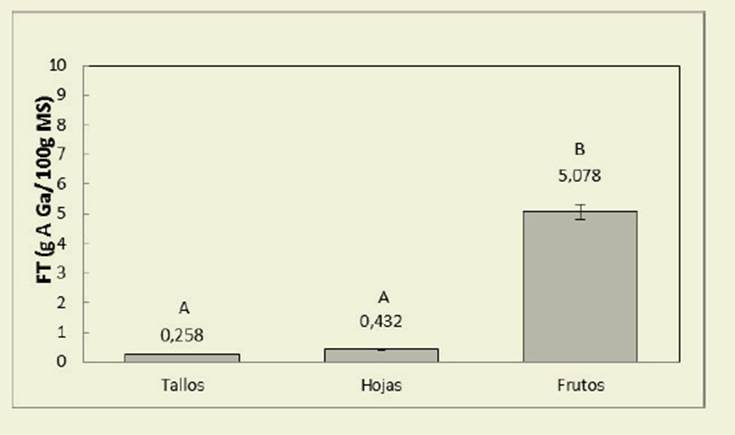
Figura 2: Contenido de fenoles totales, expresado como g AGa/100 g MS, en tallos, hojas y frutos de C. hassleri. Los valores son promedios, independientemente del momento del ciclo productivo y la condición de luz (p< 0,05) Figure 2: Content of total phenols, expressed as g AGa / 100 g DM, in stems, leaves and fruits of C. hassleri. The values are averages, regardless of the time of the production cycle and the light condition (p <0.05)
En cuanto a TT, se observó interacción entre los factores órgano y condición de luz ( Tabla 3).
Tabla 3: Contenido de taninos totales (mg AGa/g MS) en interacción órgano x condición de luz. Table 3: Content of total tannins (mg AGA/g DM) in interaction organ x light condition
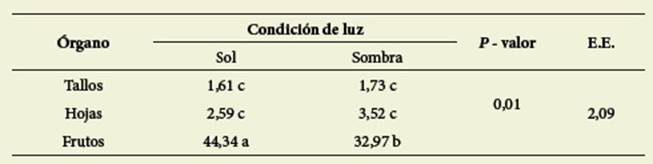
Letras diferentes indican diferencias significativas (valor p< 0,05)
Different letters indicate significant differences (p value < 0,05)
Las mayores concentracio nes se encontraron en los frutos en Csol. También hubo interacción significativa (p= 0,0023) entre los factores órgano x momento del ciclo ( Tabla 4).
Tabla 4: Contenido de taninos totales (mg AGa/g MS) en interacción órgano x momento del período de crecimiento. Table 4: Content of total tannins (mg AGa/g DM) in interaction organ x moment of growth period
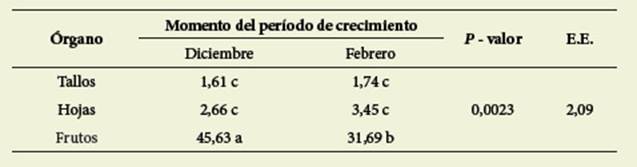
Letras diferentes indican diferencias significativas (valor p < 0,05)
Different letters indicate significant differences (p value < 0,05)
Los TT en los frutos son significativamente ma yores que en los demás órganos, con el máximo valor en los frutos en diciembre.
Respecto de los TC extraíbles, solo el factor órgano produjo diferencias signifi cativas (valor p< 0,0001) ( Figura 3).
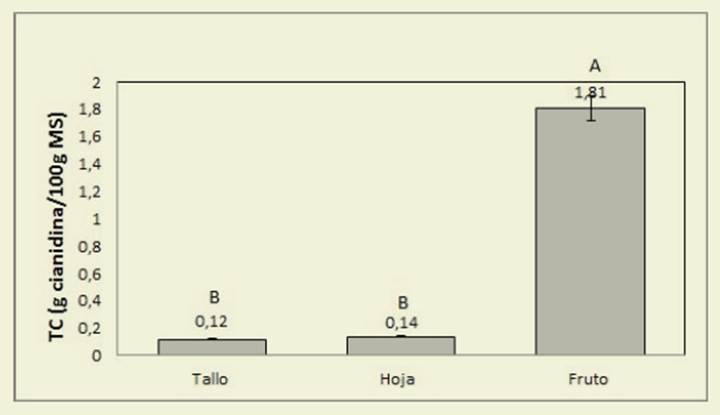
Figura 3: Contenido de taninos condensados (g cianidina/100 g MS) para tallos, hojas y frutos de C. hassleri. Los valores son promedios, sin considerar el momento del ciclo ni las condicio nes de luz (p<0,0001) Figure 3: Content of condensated tannins (g cianidine/100g DM) for stems, leaves and fruits of C. hassleri. The values are mean values, regardless the moment of the growth cycle and light conditions (p<0,0001)
Las condiciones lumínicas y el momento del ciclo no produjeron diferencias significa tivas (p=0,2242 y 0,7378 respectivamen te). El contenido de TC presentó valores medios de 0,66 y 0,72 g de cianidina/100 g de MS para las Csol y Csombra respec tivamente. Al considerar el momento del ciclo de crecimiento, los valores prome dio fueron 0,70 y 0,68 g de cianidina/100 g de MS para los meses de diciembre y febrero respectivamente.
En el contenido de flavonoides, la in teracción órgano x condición de luz ( Ta bla 5) produjo diferencias significativas (p=0,0483) encontrándose mayor con centración en las hojas de plantas creci das en sombra.
Tabla 5: Contenido de flavonoides (mg Q/g MS) en interacción órgano x condición de luz Table 5: Content of flavonoids (mgQ/g DM) in interaction organ x light condition
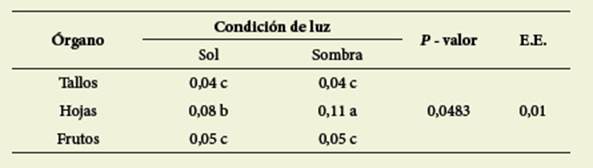
Letras diferentes indican diferencias significativas (valor p < 0,05)
Different letters indicate significant differences (p value < 0,05)
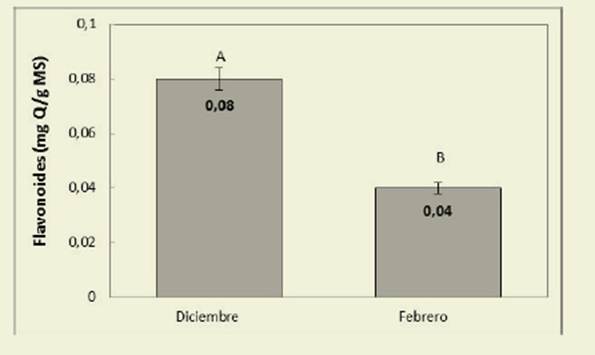
También el momento del ciclo produjo diferencias significativas (valor p <0,0001) con valores promedio de 0,08 y 0,04 mg de quercetina/gMS para diciembre y febrero respectivamen te ( Figura 4).
Discusión
La mayor producción de MV y MS ob servada en las parcelas bajo cielo abierto coincide con lo informado por Obispo et al. (2008) en Panicum y fue atribuido a una mayor actividad fotosintética en plantas expuestas al sol. Estos resultados sugieren que la presencia de C. hassleri, preferentemente debajo de los árboles, podría deberse a una mayor disponibili dad de agua en el suelo y no a un menor requerimiento lumínico. De acuerdo con Wilson & Ludlow (1991), la sustancial disminución de la temperatura en la su perficie del suelo producida por el som breo podría ser importante para mejorar el contenido de agua en el suelo, favore ciendo así la supervivencia y crecimien to de las plantas. La mayor producción de material vegetal observada en febrero ( Tabla 1) es resultante de un mayor cre cimiento acumulado al avanzar la esta ción de crecimiento, en coincidencia con lo reportado por Sosa Rubio et al . (2008) , en algunas especies poáceas y legumino sas. Valles et al . (1992 ) publicaron valo res de producción promedio de MS del orden de 1410 kg MS/Ha para otras legu minosas como Centrosema sp ., Arachis pintoi, Pueraria phaceoloides y Desmo dium ovalifolium, mientras que Sánchez et al. (2003) obtuvieron promedios de producción de MS para Cenchrus cilia ris de 1605 kg MS/ ha, comparables a los observados para C. hassleri. La calidad nutritiva en términos de PB disminuyó al avanzar el ciclo productivo ( Tabla 2), en coincidencia con lo observado por Chacón-Hernández & Vargas-Rodrí guez (2009) en Pennisetum, quienes lo atribuyeron a una reducción de la activi dad metabólica del vegetal. En este caso, el mayor porcentaje de PB en diciembre estuvo asociado a la mayor proporción de hojas en esa época ( Figura 1). El con tenido de PB, al igual que lo reportado para otras especies ( Obispo et al ., 2008 ; Piñeros et al., 2011) no fue modificado por las condiciones lumínicas. Basiga lup (2014) reportó valores promedio de 16% de PB en heno de alfalfa, similares a los obtenidos para C. hassleri, siendo los valores ideales 18 a 22% PB según este autor. Según Sosa Rubio et al. (2004), es pecies con valores de PB inferiores a un 8% de PB se consideran deficientes si se tiene en cuenta que las vacas lactantes presentan requerimientos de 9,7% de PB. De acuerdo a esto, C. hassleri podría ser tenida en cuenta como una forrajera con buen contenido proteico.
El aumento de fibra indicado por los mayores valores de FDN y FDN al final del ciclo ( Tabla 2) también ha sido ob servado en otras especies y está asociado con un incremento en los componentes de la pared celular y una disminución del contenido celular ( Chacón-Hernán dez & Vargas-Rodríguez, 2009). Valores muy similares han sido obtenidos por González et al. (2008), en alfalfa y por Kyriazopoulos et al . (2012 ) en Trifolium subterraneum. Según Basigalup (2014), los valores ideales de FDN varían entre 40 y 47% FDN y entre 32 y 36% de FDA en heno de alfalfa. Los valores obtenidos en C. hassleri se encuentran dentro de estos rangos.
En relación a los FT ( Figura 2), se ci tan para algunas leguminosas arbustivas como Tadehagi spp. y Cratylia argentea, valores muy variables de 18,3% y 0,9% respectivamente ( Lascano, 1995)
Los valores de TT ( Tablas 3 y 4), mayores para diciembre en relación a febrero podrían deberse a una mayor condensación de fenoles de alto peso molecular, producida a medida que au menta la edad de la planta, para formar taninos condensados. Los frutos presen tan en general mayores niveles de TT que suelen concentrarse en la piel ( Taiz & Zeiger, 2006) y algunas especies incre mentan su contenido en luz ( Rugna et al. 2007), aunque esto último no ha sido ob servado en nuestro caso. Con respecto a la fracción de TC, Schofield et al. (2001) informaron valores mayores de TC para otras leguminosas como Desmodium ovalifolium, Gliricidia sepium y Manihot esculenta comparados con los obtenidos para C. hassleri, aun considerando el contenido correspondiente a los frutos, en los que se observó la mayor concen tración, alcanzando un promedio de 1,81 g de cianidina/100 gMS ( Figura 3). Los TC en concentraciones superiores a los 40-50 g/kg MS en forrajes, disminuyen la digestibilidad proteica y de la MS en el rumen, sin embargo, en concentracio nes más bajas (entre 10 y 30 g/kg MS) se consideran beneficiosos debido a su efecto en la degradación ruminal de las proteínas, de modo que especies con me nos de 30 g/kg MS podrían ser incluidas en la dieta para rumiantes ( McMahon et al., 2000). De acuerdo a esto, C. hassleri podría considerarse una especie adecua da para su uso como forrajera debido a su bajo contenido en taninos condensa dos. El mayor contenido de flavonoides fue observado en las hojas y disminuyó al avanzar el ciclo ( Figura 4), lo que po dría estar relacionado con la menor pro porción de hojas al final del ciclo y con la transformación de los flavonoides con la edad.
Conclusiones
Los resultados indican que esta espe cie presenta una composición química nutricionalmente buena y sugieren que podría utilizarse como una alternativa complementaria para la alimentación de rumiantes en ambientes como los de nuestra provincia, donde no abundan los recursos forrajeros con contenidos de proteína similares a los de la alfalfa.

