











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Pappophorum Schreber es un género de gramíneas nativo del continente ameri cano con 10 especies, de las cuales 7 ha bitan en zonas áridas y semiáridas de Ar gentina y regiones limítrofes ( Pensiero, 1986). En Argentina, más específicamen te en los pastizales del sur de la provincia fitogeográfica del Monte, P. vaginatum Buckley es prácticamente la única espe cie que provee forraje para los animales durante el verano (Giorgetti, com. pers., Chacra Experimental Patagones, Car men de Patagones, Pcia. Buenos Aires, 2005). Giorgetti et al. (1997, 1998, 1999, 2000a, b) informaron que esta especie es la gramínea perenne C 4 nativa palatable más abundante durante la primavera y el verano. Esta situación la expone al so brepastoreo y actualmente se la considera como una especie decreciente ( Torres et al. 2013). En Argentina, P. vaginatum también puede ser hallada en las provin cias fitogeográficas Pampeana, del Espi nal y sudeste de la Chaqueña ( Pensiero, 1986).
Pappophorum vaginatum es una es pecie diploide con 2n=60 cromosomas ( Gould, 1966; Reeder & Singh, 1968). Garner et al. (2006), Smith (2010) y Smith et al. (2010) informaron que esta especie sería autógama o apomíctica, aunque advirtieron que se deberían rea lizar estudios más específicos sobre el sistema de reproducción de la misma, en base a sus experiencias en la obtención de cultivares de P. vaginatum en Esta dos Unidos. Por su parte, Campbell et al. (1983), Rossengurt (1984) y Pensiero (1986) informaron presencia de cleisto gamia en Pappophorum spp., mecanismo que favorece la autofecundación. Dicha especie se desarrolla en regiones áridas-semiáridas. En general, ambientes que no son óptimos para el crecimiento y la reproducción favorecen la cleistogamia ( Campbell et al., 1983).
Las especies forrajeras, nativas o na turalizadas están adaptadas a las condi ciones específicas del ambiente ( Pistora le et al., 2008). Debido a esto, la mejora genética debería orientarse a lograr la combinación de los efectos de la selec ción natural en el ambiente local y los de la selección artificial para caracteres agronómicos, especialmente los vincu lados al control de la implantación y la producción de forraje y semillas ( Muji ca, 2010a, b). En este sentido, además de conocer aspectos agronómicos de la es pecie, es necesario conocer las propieda des genéticas de la población en estudio ( Abbot & Pistorale, 2010). Estas propie dades están determinadas por la magni tud relativa de los componentes de la va rianza ( Falconer & Mackay, 1996). Así, el cociente entre las varianzas genotípica y fenotípica se denomina heredabilidad en sentido amplio y expresa el grado en el que los fenotipos de los individuos están determinados por sus genotipos ( Han son, 1963; Falconer & Mackay, 1996; Cubero, 2013). Dicho parámetro es una de las propiedades más importantes de los caracteres cuantitativos ( Falconer & Mackay, 1996; Cubero, 2013) y puede va riar para distintos caracteres del mismo organismo, para un mismo carácter en organismos distintos, para un mismo ca rácter en poblaciones distintas del mis mo organismo ( Abbot & Pistorale, 2010) e incluso para el mismo carácter y po blación en ambientes distintos ( Falconer & Mackay, 1996; Cubero, 2013). Varios autores ( Hanson, 1963; Nyquist, 1991; Falconer & Mackay, 1996; Cubero, 2013) expresan que el aspecto más destacable de la heredabilidad es que permite pre decir la respuesta a la selección. Dicha respuesta, también denominada avance genético, se define como la diferencia entre los valores fenotípicos medios de los hijos de los padres seleccionados y el de la población parental antes de ser seleccionada ( Falconer & Mackay, 1996). Además, Holland et al. (2003) destacan su importancia para comparar entre di ferentes estrategias de selección.
Otro parámetro útil para determinar las posibilidades de obtener respuesta a la selección es la relación entre los coefi cientes de variación genético y ambiental ( Vencovsky, 1987). Estudios referidos a parámetros genéticos de diversos ca racteres se han realizado en varias gra míneas forrajeras desde los años 1950. Se han efectuado estudios, por ejemplo, en Bromus spp. ( McDonald et al., 1952; Casler, 1998; Aulicino & Arturi, 2002; Abbot & Pistorale, 2010), Dactylis glo merata ( Kalton et al., 1952; Casler, 1998; 2005), Festuca arundinacea ( Burton & De Vane, 1953), Paspalum dilatatum ( García et al., 2001, 2002), Thinopyrum ponticum ( Pistorale et al., 2008), Trichlo ris crinita ( Andrés & Quiroga, 2010) y en Phalaris aquatica ( Spara et al., 2014). Los objetivos de este estudio fueron de terminar: (a) los componentes de la va rianza y coeficientes de variación (a nivel fenotípico, genotípico y ambiental), y (b) la heredabilidad en sentido amplio para caracteres de interés agronómico en cua tro poblaciones espontáneas (P1, P2, P3, y P4) de P. vaginatum recolectadas en la región semiárida de la provincia de Bue nos Aires.
Material y Método
Sitios de recolección de las semillas
Semillas de P. vaginatum se recolecta ron de cuatro poblaciones espontáneas de la región semiárida de la provincia de Buenos Aires en diciembre de 2012 ( Figura 1).
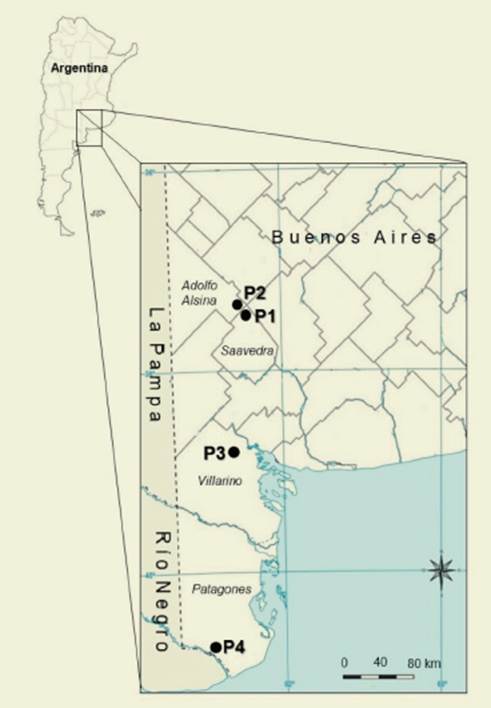
Figura 1: En el detalle del mapa de Argentina se indican las áreas donde se recolectaron las semillas de las cuatro poblaciones (P1, P2, P3 y P4) de Pappophorum vaginatum. Figure 1: The detail of the map of Argentina shows the areas where the seeds of the four populations (P1, P2, P3 and P4) of Pappophorum vaginatum were collected
Las ubicaciones geográficas de los sitios de recolección se describen en la Tabla 1.
Tabla 1: Partido y coordenadas geográficas de los sitios de recolección de las semillas de las poblaciones de P. vaginatum. Table 1: District and geographic coordinates of seed collection sites of P. vaginatum populations.
En la zona de recolección de P1 y P2 la precipitación media anual histórica (1911 a 2011) es de 665,1 mm. La temperatura media anual es de 14,9 °C y las medias anuales máximas y mí nimas son de 21,3 °C y 8 °C. Las tem peraturas máximas y mínimas absolutas son 42,5 °C (enero) y -12 °C (julio). La humedad relativa media anual histórica (1962 a 2011) es 66,25% ( INTA, 2012). El relieve es típicamente una llanura con cobertura de estrato herbáceo. En la zona de recolección de P3 la precipitación media anual histórica (1959 a 2010) es de 648,5 mm. La temperatura media anual es de 15,4 °C y las medias anuales máximas y mínimas son de 22,8 °C y 9 °C. Las temperaturas máximas y mínimas absolutas son 43,8 °C (enero) y -11,8 °C (julio). La humedad relativa media anual histórica (1960 a 2014) es 63,7% ( Servicio Meteorológico Nacio nal: SMN, 2017). El relieve es una llanura ondulada con lomas arenosas con cober tura de estrato herbáceo y comunidades arbustivas secundarias. En la zona de recolección de P4 la precipitación me dia anual histórica (1981 a 2016) es de 434,2 mm. La temperatura media anual es de 14,1 °C y las medias anuales máxi mas y mínimas son de 20,9 °C y 7,4 °C. Las temperaturas máximas y mínimas absolutas son 42,1 °C (enero) y -8,6 °C (julio). La humedad relativa media anual histórica (1981 a 2016) es 65,5% (Mi nisterio de Agroindustrias, Provincia de Buenos Aires: MA-BA, 2016). El relieve es típicamente una llanura con cobertu ra de estrato arbustivo abierto y estrato herbáceo.
Procedimientos
El 23/08/2013, se sembraron en bandejas plásticas de 128 celdas (5 cm de altura; 28 cm de ancho; 54 cm de largo), con sustrato mezcla de suelo de la Chacra Ex perimental Patagones MA-BA con arena (relación 3:1 v/v) 50 cariópsides con sus envolturas (glumelas: lemma y pálea) de cada uno de 30 genotipos diferentes de cada población (50 cariópsides/genotipo x 30 genotipos/población x 4 poblacio nes= 6000 cariópsides). El volumen de cada celda fue de 24 cm 3. El 27/09/2013, 12 plantas de cada uno de 9 genotipos (líneas puras) de cada población, obteni das luego del estudio de emergencia, se trasplantaron a bandejas plásticas de 72 celdas (5 cm de altura; 28 cm de ancho; 54 cm de largo) con sustrato mezcla de suelo de la Chacra Experimental Patago nes MA-BA con arena (relación 3:1 v/v). Cada celda tuvo un volumen de 55 cm 3. Esto fue conducido a la intemperie en La Plata (34°56’13’’ S, 57°56’41’’ O) en con diciones semicontroladas (sin incidencia de las precipitaciones) manteniendo rie go para evitar déficit hídrico (las bande jas plásticas se registraron diariamente y el riego se aplicó procurando que el volumen de tierra de cada celda estu viera siempre saturado de humedad). El riego se detuvo cada vez que comenzó a salir agua gravitacional del fondo de la celda. La humedad (%) y temperatura medias, y temperaturas medias máxi mas y mínimas del aire durante este pe ríodo realizado en La Plata (23/8/2013- 25/10/2013) fueron 81,8 % ± 0,6; 13,2 °C ± 0,6; 18,7 °C ± 0,6 y 8,2 °C ± 0,6; res pectivamente ( FCA y F-EEJH, 2013). El 25/10/2013 las 6 plantas de cada uno de los 9 genotipos (líneas puras) de cada población (6 plantas/repetición/genoti po/población x 9 genotipos/población x 4 poblaciones x 2 repeticiones/genotipo/ población= 432 plantas) se trasplantaron a un ensayo de campo (0,5 m entre geno tipos/filas x 0,25 m entre plantas dentro de cada fila) con un diseño experimental de bloques completos al azar (n=2) en la Chacra Experimental Patagones MA-BA (40° 39´ S; 62° 54´O) ( Figura 2).

Figura 2: Detalle del diseño del ensayo de campo realizado en la Chacra Experimental Patago nes. En cada bloque (bloque I, bloque II) se muestra la distribución de las plantas en cada una de las 4 poblaciones (P1 a P4) sorteadas al azar. Dentro de cada bloque, hubo 6 columnas y 9 filas por parcela (54 plantas por parcela). Alrededor de todas las parcelas se mantuvo una zona de 1 m sin vegetación durante el estudio. Su objetivo fue mantener dicha zona sin competencia radical desde las plantas allí existentes. Las malezas fueron periódicamente removidas de dicha zona. Figure 2: Detail of the design of the field trial carried out at the Patagones Experimental Farm. In each block (block I, block II) the distribution of plants in each of the 4 populations (P1 to P4) drawn at random is shown. Within each block, there were 6 columns and 9 rows per plot (54 plants per plot). A 1 m zone without vegetation was maintained around all plots during the study. The objective was to maintain this zone without root competition from the existing plants. Weeds were periodically removed from this zone.
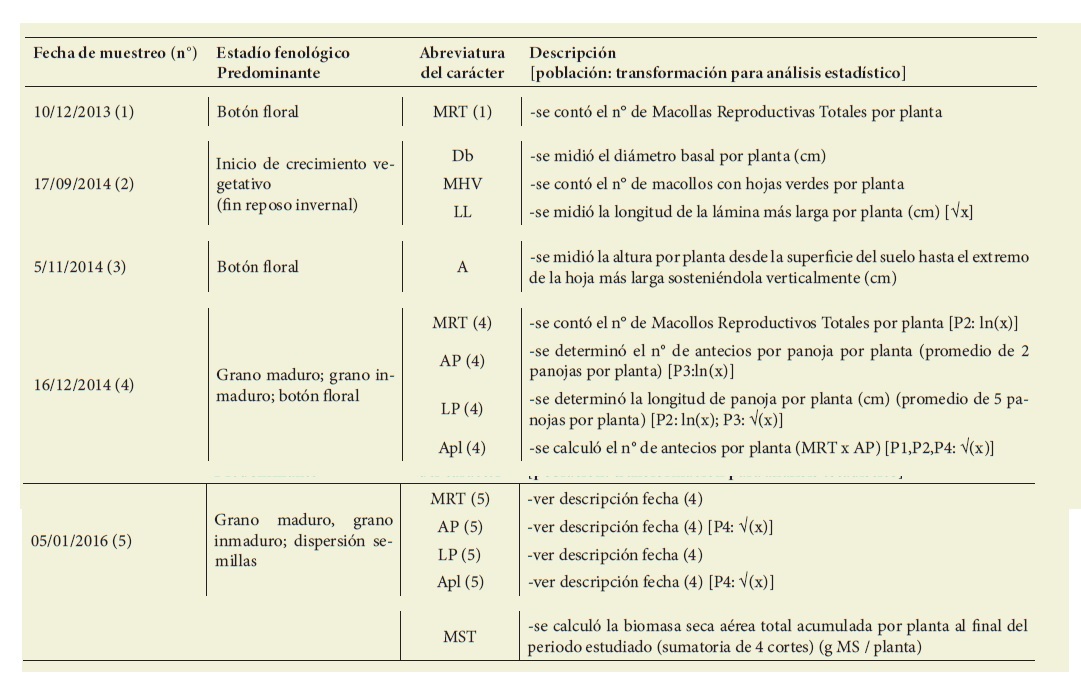
Para asegurar la supervivencia de las plantas se aplicó un riego (hasta inundar cada surco) al inicio de cada mes (diciembre, enero febrero) durante el primer verano. El estudio finalizó el 06/06/2016 (reposo invernal luego de los estudios efectua dos desde la implantación). Durante el período estudiado se registraron y determinaron caracteres vinculados a la implantación y crecimiento inicial, y a la producción de biomasa aérea y semi llas. Para el estudio de la producción de biomasa seca aérea se realizaron cuatro cortes (C1: 25/5/2015; C2: 26/11/2015; C3: 29/02/2016; C4: 06/06/2016) duran te el primer período de aprovechamien to después de la implantación. En cada momento de corte se cortaron todas las plantas. Cada planta se cortó a 5 cm de altura, se colocó en sobre de papel, se secó en estufa (6 días a 60 °C) y se pesó (g MS/planta). En la Tabla 2 se presen tan detalles de las fechas de muestreo y de los caracteres estudiados.
Tabla 2: Detalle de las fechas de muestreo y de los caracteres estudiados. Las abreviaturas indican lo siguiente: MRT: Macollas Reproductivas Totales; Db: Diámetro Basal; MHV: Macollas con Hojas Verdes; LL: Longitud de la Lámina; A: Altura; AP: Antecios por Panoja; Apl: Antecios por planta; MST: Materia Seca aérea Total. Table 2: Details of sampling dates and characters studied. The abbreviations indicate the following:= MRT: Total Reproductive Macollas; Db: Basal Diameter; MHV: Macollas with Green Leaves; LL= : Leaf blade length; A: Height; AP: Antecios per panicle; Apl: Antecios per plant; MST: Total aerial dry matter.
Análisis estadístico y genético
Los datos se analizaron mediante ANO VA para cada población en particular lo cual permitió detectar la variabilidad entre los genotipos dentro de cada una de ellas y estimar los parámetros genéti cos para cada población ( Andrés & Qui roga, 2010). Previo al análisis, algunas variables en algunas poblaciones fueron transformadas a fin de cumplir con los supuestos de normalidad y homocedas ticidad ( Sokal & Rohlf, 1984) (ver deta lle en Tabla 2).
Los componentes de la varianza fueron estimados mediante los cuadrados medios del análisis de varian za ( Burton & DeVane, 1953; Singh et al., 1993; Falconer & Mackay, 1996; Cubero, 2013). Así, la varianza ambiental, la va rianza genotípica y la varianza fenotípica se estimaron según las siguientes ecua ciones:
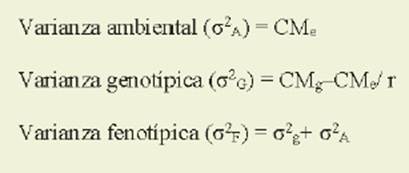
donde: CM e= cuadrado medio del error experimental; CM g= cuadrado medio de los genotipos; y r = número de repeticio nes.
Los coeficientes de variación genético, fenotípico y ambiental se calcularon se gún:

Con las estimaciones de los compo nentes de la varianza se calculó la here dabilidad en sentido amplio ( H 2) ( Bur ton & DeVane, 1953; Singh et al., 1993; Falconer & Mackay, 1996; Cubero, 2013) según la siguiente expresión:
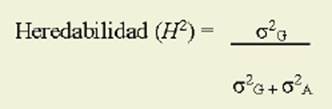
Como la varianza genética contiene los efectos aditivos y no aditivos, de bido a posibles errores de muestreo se obtuvieron algunos componentes de la varianza negativos. Debido a esto, estas estimaciones negativas se equipararon a cero al hacer sus relaciones ( Robinson et al., 1955; Aulicino & Arturi, 2002). Los análisis estadísticos se realizaron usando el programa Statistica 7.1 ( StatSoft, Inc, 2005).
Resultados
Los resultados de los análisis de la va rianza de los cuales se estimaron los pa rámetros genéticos para cada carácter en las cuatro poblaciones se muestran en la Tabla 3.
Tabla 3: Análisis de la varianza para caracteres de interés agronómico en cuatro poblaciones de Pappophorum vaginatum. CM= cuadrado medio. Los caracteres estudiados fueron: MRT: Macollas Reproductivas Totales; Db: Diámetro Basal; MHV: Macollas con Hojas Verdes; LL: Longitud de la Lámina; A: Altura; AP: Antecios por Panoja; Apl: Antecios por planta; MST: Materia Seca aérea Total. Table 3: Analysis of variance for characters of agronomic interest in four populations of Pappophorum vaginatum. CM= mean square. The characters studied were: MRT: Total reproductive tillers; Db: Basal diameter; MHV: Macollas with green leaves; LL= : Leaf blade length; A: Height; AP: Antecios per panicle; Apl: Antecios per plant; MST: Total aerial dry matter.
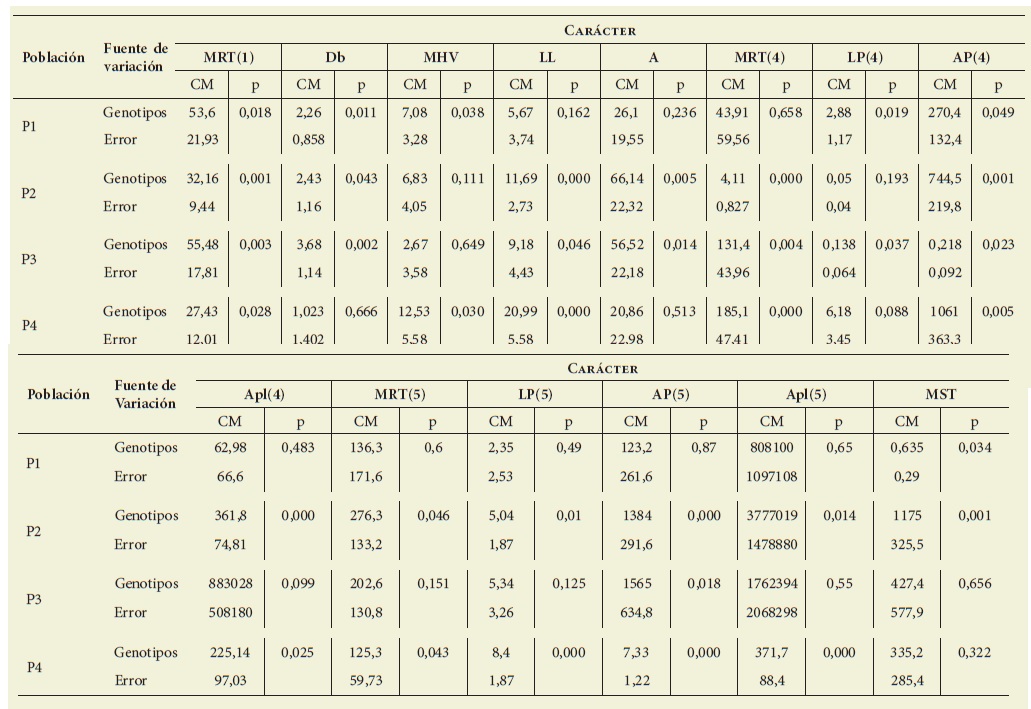
Para P1, P2, P3 y P4 hubo di ferencias significativas (p≤0,05) en 6, 12, 8 y 10 de los 14 casos (carácter-fecha de muestreo) estudiados, respectivamen te ( Tabla 3).
Por su parte en la Tabla 4 se muestran los valores de varianza ge notípica, fenotípica y ambiental con sus respectivos coeficientes de variación, la relación entre el coeficiente de variación genético y el coeficiente de variación ambiental, y la heredabilidad en sentido amplio.
Tabla 4: Varianza genética (σ 2 G), ambiental (σ 2 A) y fenotípica (σ 2 F), heredabilidad en sentido amplio (H 2) y coeficientes de variación genético (CV G) y ambiental (CV A) en caracteres de interés agronómico (carácter [fecha nro.]) en cuatro poblaciones de Pappophorum vaginatum. Las abreviaturas indican lo siguiente: MRT: Macollas Reproductivas Totales; Db: Diámetro Basal; MHV: Macollas con Hojas Verdes; LL: Longitud de la Lámina; A: Altura; AP: Antecios por Panoja; Apl: Antecios por planta; MST: Materia Seca aérea Total. Table 4: Genetic (σ2G), environmental (σ2A) and phenotypic (σ2F) variance, broad sense heritability (H2) and coefficients of genetic (CVG) and environmental variation (CVA) in characters of agronomic interest (character [date no.]) in four populations of Pappophorum vaginatum. Abbreviations indicate the following= : MRT: Total Reproductive Macollas; Db: Basal Diameter; MHV: Macollas with Green Leaves; LL= : Leaf blade length; A: Height; AP: Antecios per panicle; Apl: Antecios per plant; MST: Total aerial Dry Matter.
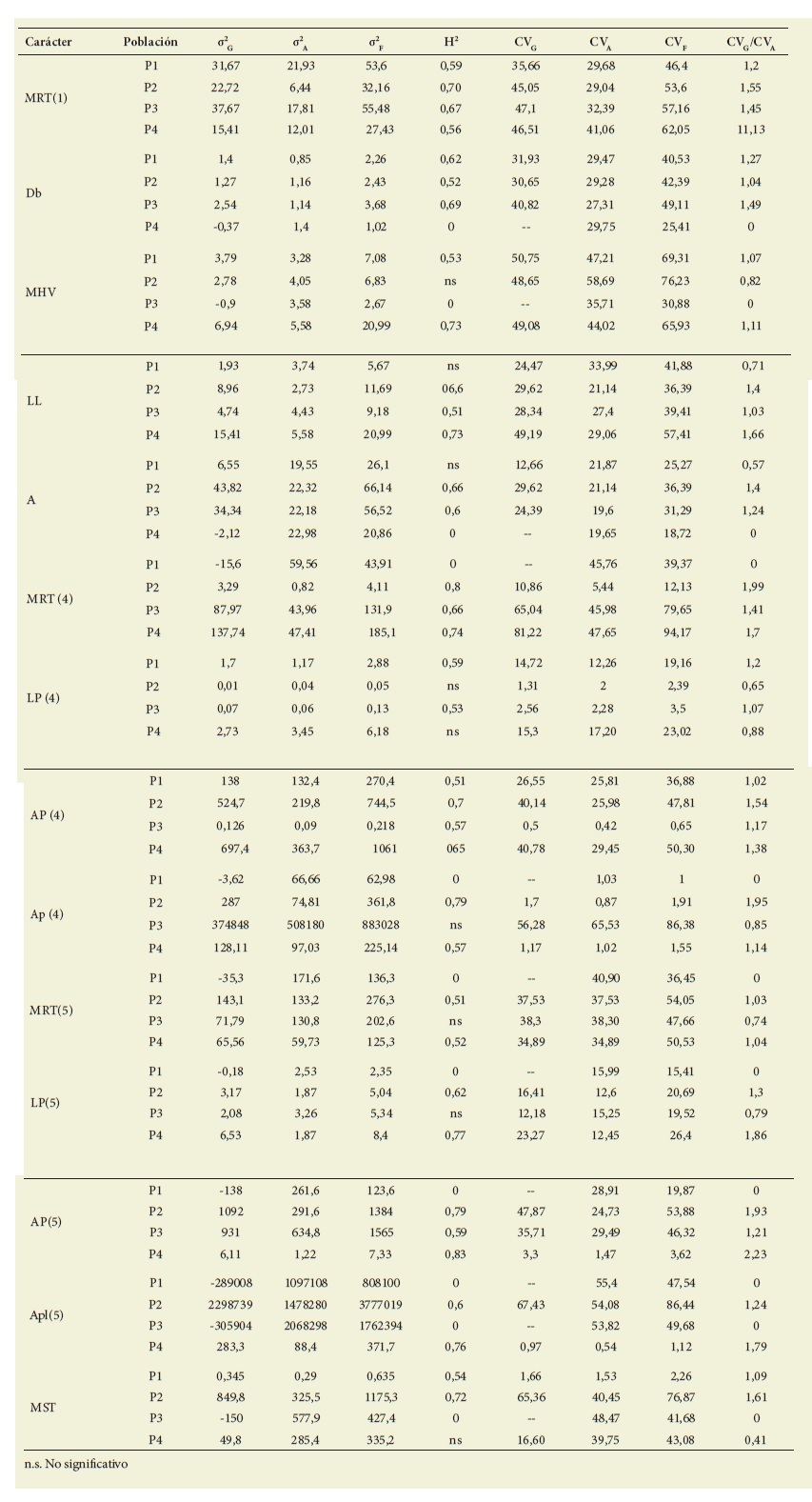
La heredabilidad en sentido amplio y la relación entre el coeficiente de variación genético y el coeficiente de variación ambiental variaron entre 0,51 y 0,83 y entre 1,02 y 2,23, respectivamente, considerando todos los caracteres en los que se pudo calcular en las cuatro pobla ciones ( Tabla 4). En 9 casos de 56 (P1: LL y A; P2: MHV y LP[4]; P3: Apl[4], MRT[5] y LP[5]; P4:LP[4] y MST), la heredabilidad en sentido amplio no se pudo calcular porque el análisis de la va rianza resultó no significativo (p>0,05) ( Tabla 3).
Discusión
La heredabilidad expresa el grado en que los fenotipos de los individuos están de terminados por sus genotipos ( Hanson, 1963; Falconer & Mackay, 1996; Cube ro 2013). Dicho parámetro es una de las propiedades más importantes de los caracteres cuantitativos ya que permite predecir el posible progreso por selec ción en un programa de mejoramiento ( Hanson, 1963; Nyquist, 1991; Falconer & Mackay, 1996 y Cubero, 2013). Los ca racteres son de alta, media y baja here dabilidad cuando presentan valores ma yores a 0,50, entre 0,20 y 0,50, y menores a 0,20, respectivamente ( Stanfield, 1971). Los resultados de esta investigación mos traron que en general los caracteres presentaron valores altos de heredabilidad en sentido amplio (entre 0,51 y 0,83), considerando las cuatro poblaciones es tudiadas ( Tabla 3). No obstante, dicha heredabilidad no se pudo calcular en 9 de 56 casos debido a que el análisis de la varianza resultó no significativo (p>0,05) ( Tabla 3). El número de macollas repro ductivas totales por planta, consideran do todas las poblaciones y fechas para las cuales se pudo calcular, tuvo una heredabilidad promedio de 0,63. Al observar por separado las fechas de muestreo (1), (4) y (5), sus valores promedio de heredabilidad fueron 0,63; 0,73 y 0,51, res pectivamente. Esto permite inferir que se puede esperar una respuesta a la selec ción para este carácter, y además mues tra que su heredabilidad se mantiene alta en los tres momentos evaluados. Otros estudios en gramíneas forrajeras tam bién informaron alta heredabilidad para el número de macollas reproductivas to tales por planta. Por ejemplo, Abbot & Pistorale (2010) en Bromus catharticus y Spara et al. (2014) en Phalaris aquatica obtuvieron valores de heredabilidad de 0,81 y 0,75, respectivamente. No obstan te, Pistorale et al. (2008) en Thinopyrum ponticum obtuvieron un valor bajo de heredabilidad (0,19) para dicho carác ter. La longitud de panoja por planta, considerando todas las fechas y pobla ciones para las cuales se pudo calcular, también tuvo una heredabilidad prome dio de 0,63. Sin embargo, se observó que en la fecha de muestreo (4) solo se pudo calcular para P1 y P3, y en la fecha de muestreo (5) para P2 y P4. Esto también permitió inferir que se puede esperar una respuesta a la selección para este ca rácter. Sin embargo, a diferencia del nú mero de macollas reproductivas totales por planta, la heredabilidad no solo que no se mantuvo alta en ambas fechas sino que en algunos casos directamente no se pudo calcular, porque el análisis de va rianza resultó no siginificativo (p>0,05). En este caso, los valores de heredabilidad de otros estudios en gramíneas forrajeras fueron variables. Por ejemplo, Abbot & Pistorale (2010) en Bromus catharticus y Spara et al. (2014) en Phalaris aquatica informaron altos valores de heredabili dad (0,66 y 0,88, respectivamente); Gar cía et al. (2001) en Paspalum dilatatum informaron valores medios (0,34), y tan to Aulicino & Arturi (2002) en Bromus catharticus como Pistorale et al. (2008) en Thinopyrum ponticum informaron va lores bajos (0,16 y 0,1, respectivamente).
El número de antecios por panoja por planta, considerando todas las pobla ciones y fechas, tuvo una heredabilidad promedio de 0,66. En este caso se podría realizar selección y esperar algún avan ce genético en cada población. Además, al igual que para el número de macollas reproductivas totales por planta, la heredabilidad se mantuvo alta en ambas fechas (es decir, [4] y [5]). En otros estu dios para este carácter se obtuvieron va lores de heredabilidad de medios a altos. Así, Abbot & Pistorale (2010) en Bromus catharticus y Spara et al. (2014) en Pha laris aquatica informaron valores altos (0,80 y 0,87, respectivamente), mientras que Aulicino & Arturi (2002) obtuvieron valores medios (0,44). El número de an tecios por planta presentó una hereda bilidad promedio de 0,68, aunque solo se pudo calcular para P2 y P4 en ambas fechas (es decir, [4] y [5]). Como resul tado, solo en dichas poblaciones se po dría esperar avance genético. De todos modos, vale aclarar que en este trabajo dicho carácter se estimó en base al nú mero de macollas reproductivas totales por planta y al número de antecios por panoja por planta.
En cuanto a los caracteres vegetativos estudiados, se observó que la heredabi lidad de la biomasa seca aérea total acu mulada por planta solo se pudo calcular para P1 y P2, y que su valor promedio fue de 0,63. Por lo tanto, se puede inferir que solo se podría esperar respuesta a la selección en las poblaciones menciona das. Este resultado coincide con lo infor mado por Pistorale et al. (2008) quienes también informaron valores altos de he redabilidad (0,55) para este carácter en Thynopirum ponticum. Casler (2005) en un estudio en Panicum virgatum tam bién informó valores altos de heredabi lidad (0,63) para dicho carácter en base a rendimientos por parcela.
Para la altura de planta también se obtuvo una heredabilidad promedio de 0,63 para las poblaciones que se pudo calcular (P2, P3). Esto coincide tanto con Andrés & Quiroga (2010) como con Casler (2005), quienes también informa ron valores altos de heredabilidad (0,9) para este carácter en Trichloris crinita y Panicum virgatum, respectivamente. Por su parte, para los caracteres diámetro ba sal, número de macollas con hojas verdes por planta y longitud de la lámina más larga por planta se obtuvieron valores promedio de heredabilidad de 0,61, 0,54 y 0,66, respectivamente para las pobla ciones que se pudo calcular (diámetro basal: P1, P2 y P4; número de macollas con hojas verdes por planta: P1 y P4; lon gitud de la lámina más larga por planta: P2, P3 y P4). Así, en general para estos tres caracteres y poblaciones las expec tativas de avance genético también son promisorias.
Otro parámetro que junto a la here dabilidad da una idea de las posibilida des de obtener algún avance genético es el coeficiente de variación genético. En este estudio, y en concordancia con la heredabilidad, se observó que en ge neral los caracteres presentaron coefi cientes de variación genéticos altos en las cuatro poblaciones estudiadas. Esto se puede inferir debido a que las rela ciones entre el coeficiente de variación genético y el ambiental fueron superio res a 1. Considerando los caracteres que también se analizaron en algunas otras investigaciones en gramíneas forrajeras los resultados coinciden parcialmente. Por ejemplo, Abbot & Pistorale (2010) informaron coeficientes de variación genéticos en Bromus catharticus de 1,66; 2,07 y 1,48 para el número de macollas reproductivas totales, número de ante cios y longitud de panoja por planta, res pectivamente. Pistorale et al. (2008) en Thynopirum ponticu informaron valores de los coeficientes de variación genéti cos de 0,48; 0,33 y 1,11, respectivamente, para el número de macollas reproducti vas totales, longitud de panoja y bioma sa seca por planta. Vencovsky (1987) y Vencovsky & Barriga (1992) también in formaron que, si la variación genética es mayor que la ambiental, la relación entre el coeficiente de variación genético y el coeficiente de variación ambiental será mayor que 1, y por ende la expectativa de respuesta a la selección será mayor en el corto plazo.
Según la población, entre 6 y 12 de los 14 casos estudiados para cada una de las poblaciones resultaron significativos (p≤0,05) en el análisis de varianza, con valores de heredabilidad altos y una re lación entre el coeficiente de variación genético y el coeficiente de variación ambiental superior a 1. Esto muestra que las expectativas de lograr algún avance genético en un programa de selección en Pappophorum vaginatum serían pro misorias para varios caracteres de interés agronómico, inclusive en el corto plazo.

