










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkLa zaranda de ideas
versão On-line ISSN 1853-1296
Zaranda ideas vol.9 no.2 Ciudad Autónoma de Buenos Aires dez. 2013
ARTÍCULO
Arqueofaunas de un espacio doméstico e implicancias para el pastoreo prehispánico en el Río Grande de San Juan
Archaeofauna from a domestic space and implications for prehispanic herding in Río Grande de San Juan
Juan Maryañski*
* ANPCyT -INAPL. Juan Maryañski es egresado de la carrera de Ciencias Antropológicas de la Facultad de Filosofía y Letras de la Universidad de Buenos Aires. Actualmente se desempeña como becario doctoral de la Agencia Nacional de Promoción Científica y Técnológica y doctorando de la Universidad de Buenos Aires. Su tema de investigación es la arqueología del pastoreo en los valles de prepuneños surandinos. E-mail: juanmki@gmail.com
Recibido: agosto de 2013
Aceptado: octubre de 2013.
RESUMEN
Este trabajo presenta el análisis de un conjunto arqueofaunístico recuperado en un espacio doméstico del sitio Chipwayko (denominado Complejo I). El mismo conforma un asentamiento conglomerado ubicado en la cuenca del Río Grande de San Juan (sudoeste de Potosí, Bolivia), ocupado durante momentos prehispánicos tardíos. A partir de este caso de estudio, se exploran aspectos relacionados con la formación del conjunto y su relación con actividades pastoriles. Se discuten algunos problemas interpretativos involucrados en la zooarqueología de espacios domésticos y se propone un abordaje basado en la información etnoarqueológica generada entre pastores surandinos. Los patrones de descarte observados en las residencias principales de pastores actuales presentan consistencias con aquellos registrados en el Complejo I, sugiriendo un rol semejante en la cadena productiva del pastoreo. Por otra parte, los resultados obtenidos indican que los patrones más amplios de producción pastoril en el área operaron bajo formas distintas a las registradas etnográficamente.
Palabras Clave: Zooarqueología; Camélidos; Pastoreo; Espacio doméstico; Período Prehispánico Tardío.
ABSTRACT
This paper presents the analysis of an archaeofaunal assemblage recovered at a domestic space from Chipiwayko site (named Complex I). Chipiwayko is an aggregated settlement occupied during late prehispanic times located at Río Grande de San Juan basin (southwestern Potosí, Bolivia). On the basis of this case study, aspects of assemblage formation and their relation with herding activities are explored. Some interpretative problems involved in zooarchaeological studies of domestic spaces are discussed and an approach based on ethnoarchaeological data of modern southandean herders is proposed. Bone discard patterns observed at modern herders main residences are consistent with that of Complex I, suggesting a similar role in the pastoral production chain. Although, these results also indicate that wider patterns of pastoral production in the area operated under different ways than those ethnographically recorded.
Key words:Zooarchaeology; Camelids; Pastoralism; Domestic space; Late Prehispanic Period.
INTRODUCCIÓN
Los estudios arqueofaunísticos de asentamientos con arquitectura remanente han sido de utilidad para abordar las prácticas económicas de las sociedades agropastoriles surandinas. Estos estudios han aprovechado la información generada a partir de muestras procedentes de ámbitos de vivienda, áreas de descarte extramuros o basureros generales (e.g. Madero 2004; Izeta y Scattolin 2006). Un problema relevante en estos casos es el de la resolución contextual de las evidencias empleadas y el alcance de las inferencias asequibles a partir de las mismas. En este sentido, los materiales recuperados en espacios domésticos son por lo general considerados como (1) representativos de un marco espacio-temporal amplio, válidos para resolver preguntas tales como la importancia relativa de ciertos recursos en un tiempo/lugar determinado; o bien (2) como punto de partida para explorar hipótesis específicamente relacionadas con el rol de dichos espacios -o sus ocupantes- en un contexto social mayor. Un ejemplo paradigmático de esto último es la búsqueda de diferencias sociales reflejadas en el acceso a los recursos (e.g. Sandefur 2001).
Si bien estos últimos enfoques aprovechan el valor de la información contextualmente específica como recurso inferencial, pocas veces se cuenta con otra fuente de expectativas más allá de los supuestos a priori sobre cómo deberían estructurarse las actividades de producción, apropiación, consumo, etc. entre los grupos que habitaron los espacios bajo estudio. El propósito de este trabajo es abordar estos problemas, enfatizando la utilidad de la información etnoarqueológica generada entre grupos pastoriles surandinos. Para esto, tomamos como base el análisis de los materiales recuperados en una antigua vivienda del sitio Chipiwayko, un extenso asentamiento conglomerado ocupado durante momentos prehispánicos tardíos (siglos X a XV d.C.). Nuestro argumento -que buscaremos defender a partir de este caso de estudio- es que el pastoreo fue una actividad central para los antiguos habitantes de Chipiwayko, siendo las viviendas el escenario donde los camélidos domésticos fueron apropiados y consumidos, operando, en algunos aspectos, de modo semejante a las viviendas pastoriles actuales.
ARQUEOFAUNAS DE ESPACIOS DOMÉSTICOS
Los restos faunísticos procedentes de ámbitos domésticos, sobre todo en asentamientos extensos e internamente diferenciados, suelen jugar un rol destacado a la hora de postular la ocurrencia de fenómenos como la especialización productiva, la centralización económica o la distribución diferencial de recursos al interior de una sociedad (ver por ejemplo de France 2009:107-122 y casos allí referidos). Bajo esta perspectiva, tales fenómenos se consideran indicadores útiles para identificar la variable injerencia de estamentos jerárquicos en los procesos económicos como contraparte de la independencia de las unidades domésticas, a cuyos miembros se atribuye el papel de productores directos. El procedimiento de análisis que se deriva de este enfoque consiste en atribuir el uso de ciertas estructuras o conjuntos arquitectónicos a segmentos sociales específicos para luego contrastar expectativas sobre las funciones productivas o el acceso a los recursos por parte de cada uno de ellos, bajo la premisa de que tales aspectos estarían "reflejados" en el pautamiento de los materiales allí presentes.
Sin embargo, existen algunos problemas que atentan contra la aplicación lineal de este tipo de razonamiento. El primero, largamente reconocido, es que el espectro de materiales recuperados en un contexto dado -en este caso, el espacio doméstico- no puede leerse como reflejo de aquel utilizado por quienes lo ocuparon. Más allá de la eventual destrucción posdepositacional, las variables trayectorias previas al descarte en distintas instancias del uso de los animales incide en los restos presentes y su pautamiento resultante (Meadow 1980).
El segundo problema, directamente relacionado con el anterior, es que los espacios de vivienda no son el equivalente material de "unidades domésticas", ya sean definidas como unidades de producción/consumo, co-residencia o parentesco. Estas dimensiones varían con cierta independencia a nivel inter e intra-social, por lo que su concurrencia o no en un entorno específico, como ha sido señalado en distintos trabajos, es una circunstancia a investigar más que un punto de partida analítico (Wilk y Rathje 1982; Allison 1999; Nielsen 2001). En función de esto, resultaría más apropiado invertir el procedimiento de estudio, abordando los espacios de vivienda y sus materiales asociados a partir del conjunto de actividades a ellos vinculados; esto es, como ámbitos arqueológicamente reconocibles donde tienen lugar al menos parte de las actividades de residencia -descanso, procesamiento y consumo de alimentos, etc.- de un grupo que no necesariamente se superpone con la categoría antropológica de unidad doméstica (Nielsen 2001). Esto permitiría, en un segundo momento, dar cuenta de las características organizativas de los actores sociales involucrados.
Desde un punto de vista etnográfico, Wilk y Netting adoptan un enfoque semejante cuando ponen el acento en la dimensión de "lo que la gente hace" o activity spheres (1984:5) por sobre las definiciones estructurales (e.g. parentesco) para aproximarse a la variabilidad empírica de los grupos domésticos. Según los autores, estos se corresponden con grupos de personas que articulan un máximo de actividades en común (e.g. co-residencia, producción y distribución de recursos), sin que ellas impliquen necesariamente a todos los miembros de dichos grupos o se circunscriban exclusivamente a los mismos. Un ejemplo de esto, usual entre campesinos surandinos actuales, es la participación regular de algunos miembros de un grupo co-residente en actividades productivas que incluyen, además, a otras personas (e.g. cosecha, molienda). Otro ejemplo, inverso pero también frecuente, es la existencia de una misma estrategia económica y de un capital común (e.g. un rebaño) en un grupo emparentado pero que ocupa más de una residencia simultáneamente (Nielsen 2001).
La ventaja de esta perspectiva es que centra el interés en las actividades domésticas -inferibles a partir de la investigación arqueológica- sin depender de la definición a priori de "unidades domésticas", entendidas como componentes abstractos de un sistema social mayor (Wilk y Rathje 1982). Los espacios de vivienda conforman una unidad de análisis arqueológica privilegiada, aunque no la única, para tal cometido. De este modo, el conocimiento sobre qué recursos animales -y cómo- se incorporan al registro arqueológico de las viviendas posee el potencial para inferir aspectos organizativos de una sociedad, siempre que se atienda a las actividades y procesos involucrados en la formación de estos conjuntos y no se los considere "reflejos" de una unidad social preestablecida.
La información etnoarqueológica producida entre pastores y agropastores del Altiplano Sur provee una vía de entrada de suma utilidad a esta cuestión. Una práctica recurrentemente documentada en estas poblaciones es la movilidad estacional que resulta en el uso de múltiples residencias a lo largo del año. Estas son usualmente clasificadas por los etnógrafos en dos grupos: (1) la casa o residencia principal, caracterizada por mayores tiempos de ocupación -incluso utilizada de forma permanente-; y (2) las estancias o puestos, que conforman asentamientos de uso temporario ligados al aprovechamiento estacional de pasturas y áreas agrícolas distantes (Tomka 1994; Yacobaccio et al. 1998; Nielsen 2000, Caracotche 2001; Göbel 2002). Desde un punto de vista material, las casas suelen presentar una mayor diversidad artefactual, extensión y complejidad arquitectónica, siendo asimismo escenarios de un espectro de actividades más amplio. Estas investigaciones coinciden en señalar que tales rasgos conforman los principales elementos para identificar arqueológicamente los espacios residenciales o de vivienda. De aquí en más, emplearemos este término en forma restrictiva, para hacer referencia a aquellos espacios construidos de uso prolongado y multipropósito (i.e. excluyendo a las instalaciones de uso temporario y/o específico).
Pero no sólo es el tiempo de permanencia lo que distingue ambas instalaciones: la casa se destaca por su centralidad en las actividades que conforman el ciclo de producción pastoril. En este sentido, los estudios etnoarqueológicos señalan que la "cadena conductual" (sensu Schiffer 1975) de la explotación pastoril tiene a la casa como locación primordial. Allí transcurren invariablemente las actividades de sacrificio y procesamiento inicial de los animales de rebaño, así como gran parte del procesamiento posterior, cocción, consumo y descarte primario de huesos. Los ritmos de matanza están normalmente dictados por las necesidades de consumo familiar y los animales de rebaño conforman el principal recurso faunístico consumido por los pastores. Aunque la caza no está ausente en las economías domésticas, la información etnográfica e histórica indica que su relevancia en las prácticas de subsistencia es, al menos desde tiempos sub-actuales, considerablemente menor (Göbel 2003:216).
El sacrificio de llamas es una actividad ritualizada que tiene lugar en sectores específicos de la residencia principal (corral, patio). Es posible que en los asentamientos conglomerados prehispánicos, dada su alta densidad edilicia, estas locaciones no presenten continuidad espacial con las áreas susceptibles de ser individualizadas como "viviendas". Sin embargo, las materialidades involucradas en el sacrificio pueden ser un buen criterio para definir su ocurrencia a nivel de sitio (Nielsen 2000; Yacobaccio y Malmierca 2006). De todas maneras, el hecho de que la matanza y ulterior procesamiento tengan lugar en las casas (en sentido amplio) resulta en la disponibilidad de animales completos -y de sus huesos- en estos espacios. Una vez procesadas, la mayoría de las unidades resultantes son almacenadas y posteriormente consumidas y descartadas allí, destinándose una porción menor al consumo inmediato luego del sacrificio. Esto es relevante para el pautamiento de las arqueofaunas allí depositadas, dado que esta secuencia de actividades conforma el principal proceso responsable de la acumulación de materiales óseos en las viviendas principales de pastores y agro-pastores actuales (Tomka 1994; Yacobaccio 1994; Yacobaccio et al. 1998).
Las investigaciones etnoarqueológicas han revelado, sin embargo, la ocurrencia de distintos procesos que involucran el descarte de restos óseos fuera del espacio doméstico. Estas prácticas han recibido particular atención en función del interés por plantear correlatos arqueofaunísticos que permitieran detectar tipos de sitios a partir de sus pautas de descarte (Tomka 1994; Yacobaccio 1994; Kuznar 1995; Yacobaccio et al. 1998; Yacobaccio y Madero 2001). Desde el punto de vista de la casa, y tomando como parámetro un esqueleto completo, esto generaría conjuntos faunísticos con "ausencias" de elementos anatómicos1 (lo que O'Connor llamó depleted [1993:64]). Estos procesos incluyen la donación ocasional de unidades de consumo a terceros; el transporte de partes -frescas o charqueadas- a los puestos de uso temporario y el descarte previo al consumo de algunas unidades con bajo contenido alimenticio. No obstante, los estudios actualísticos mostraron una importante variación en las unidades anatómicas involucradas en estos procesos. Esto implica que, salvo por el eventual desecho pre-consumo de algunas partes, no es esperable un patrón discernible al respecto en conjuntos agregados (Yacobaccio y Madero 2001).
CHIPIWAYKO Y EL COMPLEJO I
Chipiwayko es un extenso sitio residencial multicomponente, emplazado a 3250 msnm sobre una terraza fluvial en la margen izquierda del río Talina. Este último es uno de los principales afluentes del Río Grande de San Juan (en adelante RGSJ, Mapa 1), alternativamente denominado San Juan del Oro -en Bolivia- o San Juan Mayo. Con una superficie de ca. 50 ha, Chipiwayko es el sitio más grande documentado hasta la fecha en la región, aunque la presencia de sectores con variable densidad edilicia así como de superposiciones constructivas sugiere que esta representa la máxima extensión de un asentamiento que creció gradualmente. Distintas fechas radiocarbónicas sitúan la historia de ocupación del sitio desde al menos el Siglo X hasta mediados del Siglo XV, siendo esta edad "tope" coherente con la ausencia de cerámica de filiación Inka en superficie y estratigrafía (Nielsen et al. 2010).
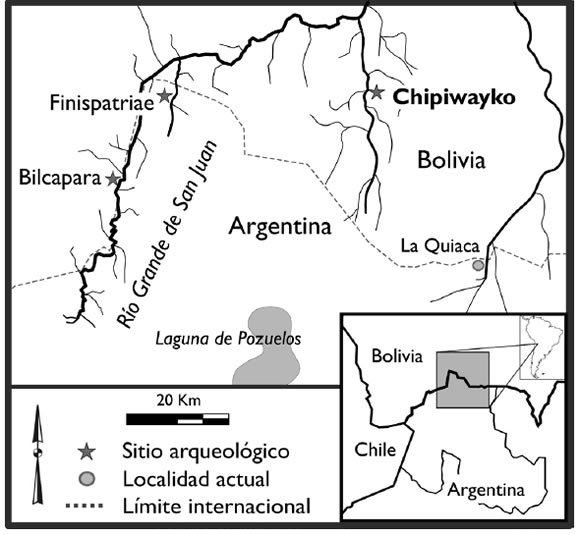
Mapa 1. Río Grande de San Juan y ubicación de Chipiwayko y de otros sitios mencionados en el texto.
El sitio está cubierto por un espeso bosque de churqui (Prosopis ferox) y cactus columnares (Echinopsis sp), lo que sumado a la densidad y pobre conservación de la arquitectura dificulta el reconocimiento de rasgos constructivos en superficie. Sin embargo, a partir de las investigaciones desarrolladas desde 2007 se logró detectar la presencia de espacios públicos, vías de circulación interna, áreas de descarte y complejos de estructuras constructivamente independientes (Nielsen et al. 2010). El término "complejo arquitectónico", aquí utilizado, refiere a un conjunto de espacios y estructuras cuya disposición material permite establecer su relación funcional (Nielsen 2006:73). Uno de ellos -denominado Complejo I (en adelante, CI)- ha sido objeto de excavaciones en área y de allí proceden los materiales aquí tratados.
Este espacio se compone de al menos ocho estructuras adosadas de planta ortogonal (Figura 1). Siete de ellas se disponen en torno a un recinto de mayores dimensiones, denominado Recinto 8. Tres de estas estructuras (Recintos 5, 6 y 7) fueron excavadas de forma total, mientras que en una cuarta (Recinto 9) se practicaron excavaciones parciales. En estas últimas se observaron restos de techumbre consistentes en paja quemada y barro (torta), situación que no comparten con el Recinto 8, también parcialmente excavado y que, por esta característica y su mayor dimensión, puede ser definido como un patio.
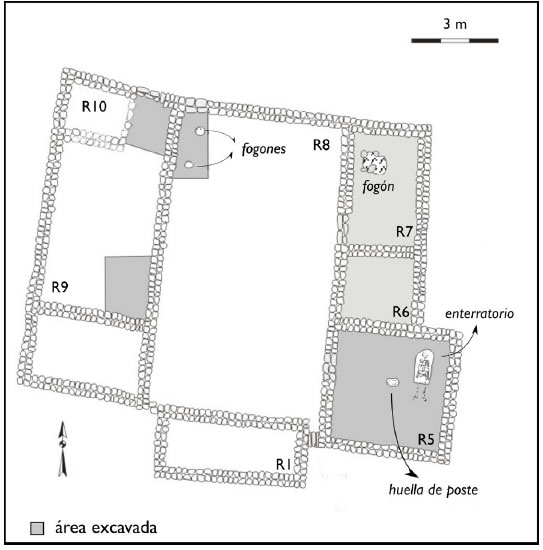
Figura 1. Planimetría del CI de Chipiwayko (modificado de Nielsen et al. 2010).
Las excavaciones revelaron un enterratorio dispuesto en una fosa en el Recinto 5, pisos empedrados con lajas en los recintos 6 y 7, un fogón formalizado en este último y fogones en cubeta en los recintos 8 y 9. La presencia de sedimentos rubefaccionados y rocas de apoyo en el fogón del Recinto 7 permite interpretar tentativamente este espacio como un ámbito de cocción de alimentos. Los materiales recuperados incluyen restos óseos, implementos de molienda, instrumentos de metal y numerosos artefactos cerámicos completos o utilizables (Nielsen et al. 2010). Estos hallazgos indican que el CI se trató de un ámbito de vivienda, cuyo diseño (recintos ortogonales en torno a un patio) ha sido observado en otros complejos y sitios del área, sugiriendo una modalidad recurrente en la región.
La mayoría del repertorio artefactual mencionado fue hallado sobre los pisos de los recintos, siendo escasa la presencia de materiales en los depósitos de sedimento eólico que rellenan las estructuras. Esto sugiere que corresponden mayormente al descarte de facto y provisorio (sensu LaMotta y Schiffer 1999) de materiales durante los momentos finales de uso del complejo (Nielsen et al. 2010). El Recinto 8 es el espacio con menor superficie excavada, aunque de allí proviene más de la mitad (n = 353) del conjunto arqueofaunístico recuperado (N = 661). Su densidad relativa y el hecho de estar acumulados contra una esquina del espacio descubierto (patio) resultan, en principio, consistentes con su carácter de desechos provisorios, i.e. basura transitoriamente depositada para su ulterior descarte definitivo.
EL REGISTRO ARQUEOFAUNÍSTICO
El conjunto del CI presenta una baja diversidad taxonómica (Tabla 1). Los restos de artiodáctilo -incluyendo camélidos e indeterminados- superan el 90% del total de los especímenes identificados y presentan abundantes evidencias de utilización humana. Los artiodáctilos indeterminados incluyen fragmentos de costillas, vértebras y huesos largos sin mayores rasgos diagnósticos que muy probablemente correspondan a camélidos. Tales materiales conforman el centro de este trabajo, sobre los cuales se analizaron variables relativas a la preservación posdepositacional, las modificaciones óseas, la frecuencia de unidades anatómicas y las edades de muerte. Estos tuvieron como fin discernir las actividades humanas involucradas en la formación del conjunto como punto de partida para su comparación con los modelos actualísticos disponibles.
Tabla 1. Abundancia taxonómica en el CI de Chipiwayko. NISP = número de especímenes identificados; EP = evidencias de procesamiento.
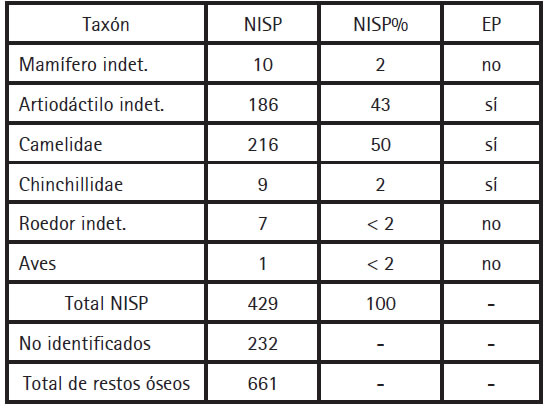
En el CI se identificaron, además, nueve restos de vizcacha (Lagidium sp) comprendidos por huesos axiales y apendiculares, de los cuales seis se encuentran fragmentados. Una tibia de vizcacha presenta escotaduras (sensu Mengoni Goñalons 2010) en su diáfisis, señal de haber sido intencionalmente fracturada, lo que sugiere fuertemente el uso humano de estos roedores. También se hallaron siete huesos apendiculares correspondientes a roedores pequeños. Estos se encuentran distribuidos en los tres recintos e invariablemente completos, sin presentar signos claros de uso humano. Un hueso largo de ave, sin evidencias de modificación antrópica, fue identificado en el Recinto 9.
Dentro de la categoría camélido se identificaron distintos grupos de tamaño a partir de criterios osteométricos, aunque sólo cumplieron los requisitos para ser medidas dos falanges segundas (atribuidas a llama-guanaco y vicuña) y una falange primera (atribuida a llama-guanaco). Este grupo presenta un importante número de restos no fusionados y algunos huesos quemados, lo que limitó significativamente la posibilidad de llevar adelante este tipo de análisis. En principio esto indica, junto a la presencia de vicuña, la posible depositación de llamas y guanacos.
Tal ambigüedad puede reducirse en este caso al considerar la información sobre las edades de muerte arrojadas por el conjunto. Para estimar el perfil de mortalidad utilizamos las evidencias de fusión de elementos poscraneales a partir del cronograma ofrecido por Kent (1982). Distinguimos tres grupos de elementos cuya fusión ocurre en rangos de edad diferentes, acotados y no superpuestos2 (Yacobaccio 2007). En este esquema, la proporción de huesos no fusionados en cada grupo permite estimar la mortalidad de ese rango etario, mientras que la de fusionados indican la supervivencia con posterioridad al mismo. En la Tabla 2 se observa que la proporción de casos fusionados es en general baja, decreciendo considerablemente en cada categoría etaria. Esto indica un ingreso predominante de animales menores a los 2-3 años en el CI, que incluye probablemente una importante cantidad de animales muertos antes del primer año de vida. Aunque se trata de una muestra pequeña, este patrón es consistente con lo registrado en otros conjuntos del área (ver más adelante). En relación con los casos de camélidos grandes taxonómicamente ambiguos, estos resultados sugieren la importancia de las formas domésticas -i.e. llamas- en el CI. Aunque no descartamos el ingreso de guanacos al mismo, la estructura etaria no concuerda con el panorama esperado en una situación de intensa captura de animales silvestres (Hesse 1982), a menos que se asuman estrategias de caza marcada y recurrentemente selectivas.
Tabla 2. Perfil de mortalidad de camélidos en el CI de Chipiwayko basado en la fusión de elementos poscraneales.
Los huesos de artiodáctilo (camélidos e indeterminados) presentan evidencias de haber estado expuestos durante cierto tiempo luego de su depositación, aunque esto no parece haber sido homogéneo para todos los especímenes. Un 48% de los mismos mostraron distintos grados de meteorización, aunque solo una menor proporción (1,3%) presentó estadios superiores a 2 (Behrensmeyer 1978). Por otra parte, en un 3% de los mismos se registró la acción de roedores. Esto sugiere un tiempo variable de exposición de los materiales y es consistente con una acumulación progresiva de los mismos, antes del colapso de la arquitectura y su subsecuente enterramiento. No obstante, no se detectó un patrón de destrucción de huesos en función de su densidad mineral ósea: el análisis de correlación entre los valores densitométricos aportados por Stahl (1999) y la frecuencia relativa de elementos presentes -medida en NISP normalizado (Grayson y Frey 2004)- para 62 pares de datos arrojó un resultado negativo (rs = -0,13) y no significativo (p = .31). Otro indicador empleado a propósito de esto fue la frecuencia de las porciones epifisiarias distales con respecto a las proximales en húmeros y tibias. Como las primeras poseen una densidad considerablemente superior (Stahl 1999), su sobre-representación puede indicar la ocurrencia de intensos procesos de destrucción no antrópica. Esta relación fue pareja en húmeros (7 a 6), mientras que en tibias fue de 1 a 2, siendo consistente con lo observado en el análisis anterior. Esto permite sostener que la eventual destrucción in situ de materiales óseos no afectó la representación de las distintas unidades anatómicas.
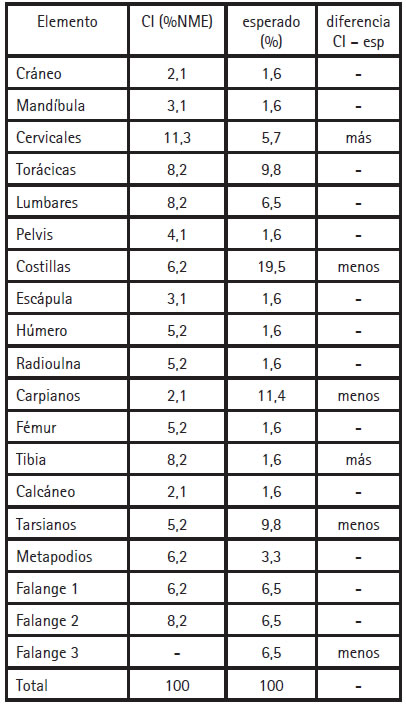
Nuestra principal hipótesis es que, al igual que en las viviendas pastoriles actuales, las llamas fueron sacrificadas, procesadas y mayormente consumidas y descartadas en el espacio de vivienda. Esto supone que la frecuencia de los diferentes elementos óseos recuperados se corresponderá, en líneas generales, con aquella en un animal completo. Ambos grupos de datos se presentan en forma porcentual en la Tabla 3. Las frecuencias para el CI fueron calculadas a partir del número mínimo de elementos (NME) definidos según zonas diagnósticas (Mengoni Goñalons 2010), obteniéndose un NME = 108. También se consignan las diferencias mayores a 4% entre ambos grupos de datos. Este es un valor arbitrario que permite distinguir los elementos cuyas frecuencias se apartan de aquella "esperada" (calculadas como porcentajes en un esqueleto completo). Como se desprende de la Tabla 3, la mayoría de los huesos fueron descartados en función de su frecuencia en un camélido completo, siendo consistente con nuestra expectativa inicial. Dentro de esta tendencia general, la sobre-representación de algunos elementos (tibia y cervicales) y la sub-representación de otros (huesos articulares, falange tercera y costillas) no puede explicarse por sus diferentes chances de preservación.
Tabla 3. Frecuencia de elementos anatómicos en el CI y en un camélido completo. Se consignan las diferencias mayores a 4%.
Una manera de evaluar esta variabilidad es considerar los recursos alimenticios asociados a cada elemento óseo. Para esto empleamos cuatro categorías propuestas por Yacobaccio y coautores (1998) que agrupan los distintos elementos en función de su variable contenido de carne y médula (Tabla 4). Estos investigadores advirtieron un patrón recurrente en los desechos óseos producidos por los pastores actuales del área de Susques (Jujuy), caracterizado por el predomino de huesos asociados a mayores contenidos de carne, el cual atribuyeron a prácticas selectivas. En la Tabla 4 volcamos por un lado las frecuencias de cada categoría en el CI y, por otro, las registradas en asentamientos pastoriles actuales y en un esqueleto completo de camélido. Puede observarse que los elementos representados en el CI se ajustan fuertemente a los patrones de descarte observados entre los pastores actuales de la Puna jujeña (Gráfico 1). Estadísticamente, ambas muestras presentan distribuciones muy semejantes (X2 = 0,99; p = .91). A su vez, la distribución de estas categorías en el CI se diferencia significativamente de aquella presente en un esqueleto completo (X2 = 12,8; p = .01). Esto sugiere la ocurrencia de procesos de formación muy semejantes en ambas situaciones, probablemente ligados al tratamiento humano de las carcasas. Al menos dentro del segmento apendicular (Grupos 2 y 3), los huesos que poseen exclusivamente médula son menos frecuentes.
Tabla 4. Frecuencia de elementos anatómicos agrupados según recursos (en porcentajes). aPromedio de sitios principales y temporarios (datos de Yacobaccio et al. 1998).
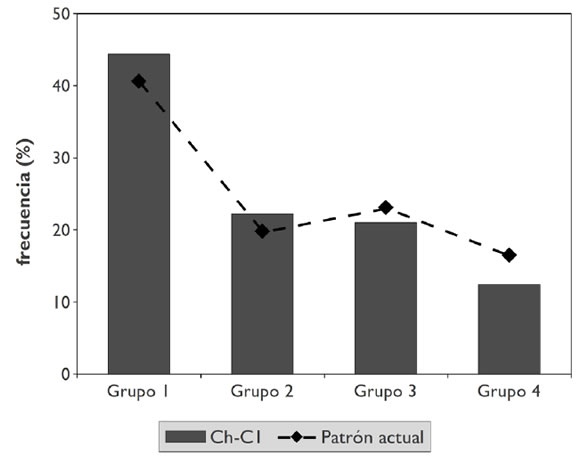
Gráfico 1.Comparación entre la frecuencia de grupos de elementos anatómicos en el CI y en sitios de pastores actuales (basada en la Tabla 4).
Esto no significa, sin embargo, que dicho recurso no haya sido aprovechado. Tomka (1994) observó que la médula ósea no es procesada de forma independiente, sino que se consume en guisos conjuntamente con la carne, fracturándose los huesos inmediatamente antes de su cocción. Esta situación da cuenta de los patrones observados y explica la presencia de negativos de impacto en un 15% (n = 24) de las diáfisis de artiodáctilos (camélidos e indeterminados). También la fragmentación de un calcáneo -sobre dos presentes- y de la totalidad de las falanges proximales (n = 14), que además presentan escotaduras y marcas de percusión. Estas últimas son unidades con escasísima carne asociada pero con cierto contenido de médula ósea (De Nigris y Mengoni Goñalons 2005) por lo que tales patrones sugieren el aprovechamiento ocasional de dicho recurso.
Las falanges segundas y el resto de los carpianos y tarsianos, en cambio, no poseen este tipo de evidencias estando invariablemente completos y en general en frecuencias muy bajas, a la vez que no se han registrado falanges terceras en el conjunto. Los carpianos y tarsianos están asociados a contenidos alimenticios mínimos, pero pueden quedar eventualmente adheridos a los huesos largos incluso durante la cocción (Yacobaccio et al. 1998). Un hueso del carpo (unciforme) presentó huellas de corte, indicando actividades de desarticulación que, sin embargo y por lo recién comentado, es difícil saber si tuvieron lugar en el espacio intramuros. Entonces, más allá de que la falta de rasgos diagnósticos mayores en ciertas unidades (e.g. costillas) incida en su sub-representación, las "ausencias" recurrentes corresponden a huesos sin recursos alimenticios asociados, lo que sugiere que siguieron trayectorias de depositación distintas luego del procesamiento inicial.
Es interesante mencionar que entre los agro-pastores de Alota-Copacabana (Norte de Lípez) se registró también una configuración semejante en los conjuntos óseos de las residencias permanentes (Tomka 1994). Allí se observó que: (1) el segmento cervical predomina dentro de los huesos axiales; (2) los huesos de las patas son más frecuentes que los de los pies; y (3) dentro de este último grupo, los metapodios son más recurrentemente descartados que las articulaciones y falanges. Estos conjuntos, cuyo patrón anatómico se asemeja al del CI (Tabla 3), son generados principalmente a partir del descarte de materiales resultantes de la matanza, procesamiento y consumo in situ de llamas por parte de los miembros del grupo residente.
DISCUSIÓN Y COMENTARIOS FINALES
Las evidencias hasta aquí tratadas sugieren fuertemente que en el CI actuaron los mismos procesos tafonómicos -en sentido amplio (Lyman 2010)- que en las casas pastoriles. Advertimos un patrón de aprovechamiento integral de camélidos completos, cuyas diferentes unidades anatómicas parecen haber sido descartadas según el ritmo impuesto por las necesidades de consumo de los antiguos habitantes del espacio doméstico. La representación de las unidades anatómicas, al analizarse desde el punto de vista de sus recursos alimenticios, es extremadamente consistente con aquella registrada en viviendas pastoriles actuales. En este sentido, puede defenderse que, al igual que en estas locaciones, el principal proceso de acumulación de materiales óseos en el CI resultó de las actividades de sacrificio, procesamiento, consumo y descarte de llamas in situ. Asimismo, el número relativamente bajo de especímenes recuperados es esperable en este tipo de locaciones, donde el uso persistente y estructurado promueve la limpieza periódica del espacio y el uso de áreas de descarte externas (e.g. Yacobaccio et al. 1998).
Lo dicho no implica descartar la ocurrencia de otras actividades involucradas en la formación del registro del CI, como el transporte de unidades a otros escenarios de actividad. Esta práctica fue registrada recurrentemente en la etnografía, aunque sus consecuencias materiales en los conjuntos faunísticos no resulta clara: es posible que carezcan de la intensidad y pautamiento suficiente para ser identificadas en conjuntos arqueológicos agregados. Otra cuestión que debe ser mencionada en relación con el CI es la presencia de camélidos silvestres. Hemos identificado la presencia de vicuña y eventualmente guanaco, lo que plantea la posibilidad de que la disponibilidad de llamas en pie concurra en cierta medida con prácticas de transporte selectivo de camélidos silvestres hacia el espacio de consumo, contribuyendo de esta manera a la configuración de un perfil anatómico sesgado. Esta hipótesis es difícil de corroborar en función de los límites existentes para la identificación específica de camélidos, pero sobre todo por la equifinalidad que presenta a priori con fenómenos como la distribución de recursos entre unidades de consumo diferentes o la dispersión de bienes de una misma unidad de consumo en espacios arquitectónicos distintos. En otras palabras, es difícil de discernir la ocurrencia de procesos de acumulación o remoción cultural, y es probable que ambos confluyan en la formación del conjunto doméstico.
En este punto es necesario marcar que, a diferencia lo que sucede con los perfiles anatómicos, la estructura etaria de los camélidos contrasta fuertemente con aquella observada a partir de los huesos descartados por pastores actuales de la Puna jujeña (Yacobaccio 2007). La información etnoarqueológica a este respecto señala el predomino general de especímenes fusionados, que alcanzan el orden del 90% en las casas para las categorías de fusión intermedia y tardía. Esto ha sido interpretado como consecuencia del sacrificio recurrente de animales adultos, propias de sistemas productivos "mixtos", i.e. orientados a la producción no especializada de fibra y carne (Yacobaccio 2007:151). Asimismo, es coherente con lo observado etnográficamente en otras comunidades pastoriles (Nielsen 2000; Göbel 2001).
Por otra parte, no sólo en el CI se ha observado un perfil de mortalidad inverso, sino también en otros contextos contemporáneos de distintos sitios conglomerados de la región. En el Gráfico 2 presentamos la información obtenida en tres densas acumulaciones de basura emplazadas en espacios extra-muros de los sitios Finispatriae (B1) y Chipiwayko (S1); del CI de Chipiwayko y de una excavación parcial efectuada en el patio de otro espacio doméstico en el sitio Bilcapara (R1). Los especímenes no fusionados oscilan entre 50-60% para la primera categoría, 62-75% para la segunda y 70-85% para la tercera, siendo altamente consistentes entre sí. Esto permite sostener que el perfil etario del CI no refleja el ingreso selectivo de determinados grupos de edad al espacio doméstico, dado que es constante entre contextos de distinta funcionalidad y resolución. En otras palabras, las acumulaciones de basura extramuros parecen captar desechos resultantes de actividades desarrolladas en distintos espacios -quizás en forma de descarte secundario-, incluyendo los ámbitos de vivienda, siendo los criterios de matanza recurrentes.
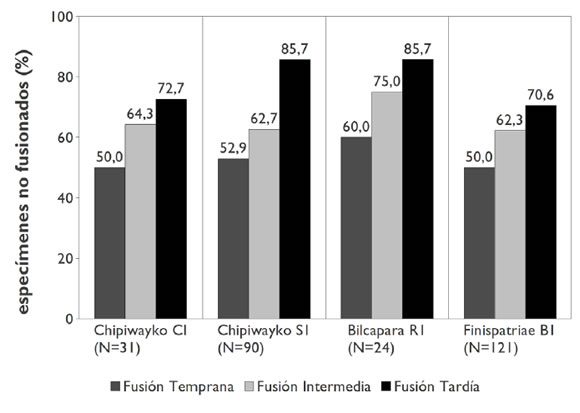
Gráfico 2.Porcentaje de especímenes no fusionados en distintos conjuntos del área, según categorías de edad.
Esto sugiere una lógica productiva distinta a la identificada entre pastores actuales del Sur Andino, ya que la conservación de animales hasta edades adultas no parece ser una estrategia preponderante. En este sentido, debe tenerse en cuenta que los estudios efectuados entre pastores actuales involucran grupos campesinos con economías pastoriles cuya producción agrícola es marginal o nula, una situación que no se corresponde con la de la cuenca del RGSJ, donde las evidencias de producción agrícola son profusas (Nielsen et al. 2013). Esto invita a considerar la incidencia de las economías agropastoriles diversificadas en las modalidades de manejo de rebaños, las cuales no incluyeron la conservación de animales hasta edades avanzadas. Asimismo, indica que aunque los camélidos una vez muertos fueron tratados de formas muy semejantes a las registradas etnográficamente, lo mismo no puede extenderse a los animales vivos.
NOTAS
1. Debe hacerse notar que las prácticas de reparto implican también el ingreso de partes de animales desde otros espacios de vivienda (Tomka 1994).
2.Para las diáfisis no fusionadas y las epífisis sueltas se contabilizó exclusivamente el número máximo entre ambas, a igual elemento y lateralidad, con el fin de no sobredimensionar la mortalidad.
AGRADECIMIENTOS
Este trabajo no hubiese sido posible sin la colaboración de los miembros del Proyecto Arqueológico Altiplano Sur (PAAS), dirigido por el Dr. Axel Nielsen, y la hospitalidad brindada por los comunarios de Chipihuayco. Los comentarios de los Dres. Axel Nielsen, Gabriel López y Matías Medina ayudaron a mejorar el manuscrito original, aunque lo aquí expresado es de mi absoluta responsabilidad.
BIBLIOGRAFÍA
1. Allison, P. 1999. Introduction. En The Archaeology of Household Activities, editado por P. Allison, pp. 1-18. Routledge, Londres. [ Links ]
2. Behrensmeyer, A. K. 1978. Taphonomic and Ecologic Information From Bone Weathering. Paleobiology 4 (2):150-162. [ Links ]
3. Caracotche, M. S. 2001. The Invisibility of Time: An Ethnoarchaeological Study of the Temporary Sites of Herders of the Southern Puna. En The Ethnoarchaeology of Andean South America: Contributions to Archaeological Method and Theory, editado por L. Kuznar, pp. 97-115. International Monographs in Prehistory, Ann Arbor. [ Links ]
4. de France, S. 2009. Zooarchaeology in Complex Societies: Political Economy, Status, and Ideology. Journal of Archaeological Research 17 (2):105-168. [ Links ]
5. De Nigris, M. y G. L. Mengoni Goñalons. 2005. The Guanaco as a Source of Meat and Fat in the Southern Andes. En The Zooarchaeology of Fats, Milks and Dairying, editado por J. Mulville y A. Outram, pp. 160-166. Oxbow, Oxford. [ Links ]
6. Göbel, B. 2001. El Ciclo Anual de la Producción Pastoril en Huáncar (Jujuy, Argentina). En El Uso de los Camélidos a Través del Tiempo, editado por G. Mengoni Goñalons, D. Olivera y H. Yacobaccio, pp. 91-115. Ediciones del Tridente, Buenos Aires. [ Links ]
7. Göbel, B. 2002. La Arquitectura del Pastoreo: Uso del Espacio y Sistema de Asentamiento en la Puna de Atacama (Susques). Estudios Atacameños 23:53-76. [ Links ]
8. Göbel, B. 2003. "La Plata no Aumenta, La Hacienda Sí": Continuidades y Cambios en la Economía Pastoril de Susques (Puna de Atacama). En Puna de Atacama. Sociedad, Economía y Frontera, compilado por A. Bendetti, pp. 199-242. Alción Editora, Córdoba. [ Links ]
9. Grayson, D. y C. Frey. 2004. Measuring Skeletal Part Representation in Archaeological Faunas. Journal of Taphonomy 2 (1):27-42. [ Links ]
10. Hesse, B. 1982. Animal Domestication and Oscillating Climates. Journal of Ethnobiology 2 (1):1-15. [ Links ]
11. Izeta, A. y M. C. Scattolin. 2006. Zooarqueología de Una Unidad Doméstica Formativa. El Caso de Loma Alta (Catamarca, Argentina). Intersecciones en Antropología 7:193-205. [ Links ]
12. Kent, J. 1982. The Domestication and Exploitation of The South American Camelids: Methods of Analysis and Their Application to Circumlacustrin Archaeological Sites in Bolivia and Peru. Tesis Doctoral inédita, Washington University, Saint Louis. [ Links ]
13. Kuznar, L. 1995. Awatimarka. Ethnoarchaeology of an Andean Herding Community. Harcourt Brace, Nueva York. [ Links ]
14. LaMotta, V. y M. B. Schiffer. 1999. Formation Processes of House Floor Assemblages. En The Archaeology of Household Activities, editado por P. Allison, pp. 19-29. Routledge, Londres. [ Links ]
15. Lyman, R. L. 2010. What Taphonomy Is, What it Isn't, and Why Taphonomists Should Care About the Difference. Journal of Taphonomy 8 (1):1-16. [ Links ]
16. Madero, C. 2004. Arqueofaunas en Sociedades Complejas: La Ganaderia Prehispánica de Camélidos en los Andes (Noroeste Argentino). En Zooarchaeology of South America, editado por G. Mengoni Goñalons, pp. 59-79. BAR International Series 1298, Archaeopress, Oxford. [ Links ]
17. Meadow, R. H. 1980. Animal Bones: Problems for the Archaeologist Together With Some Possible Solutions. Paléorient 6:65-77. [ Links ]
18. Mengoni Goñalons, G. L. 2010. Zooarqueología en la Práctica: Algunos Temas Metodológicos. Xama 23:83-113. [ Links ]
19. Nielsen, A. 2000. Andean Caravans: An Ethnoarchaeology. Tesis Doctoral inédita, Department of Anthropology, University of Arizona, Tucson. [ Links ]
20. Nielsen, A. 2001. Evolución del Espacio Doméstico en el Norte de Lípez (Potosí, Bolivia): ca. 900-1700 DC. Estudios Atacameños 21:41-61. [ Links ]
21. Nielsen, A. 2006. Plazas para los antepasados: Descentralización y Poder Corporativo en las Formaciones Políticas Pre-incaicas de los Andes Circumpuneños. Estudios Atacameños 31:63-89. [ Links ]
22. Nielsen, A., C. Angiorama, J. Maryañski, L. López y F. Ávila. 2013. La Organización del Espacio en la Sub-región San Juan Mayo Durante el Período de Desarrollos Regionales. Trabajo presentado en el XVIII Congreso Nacional de Arqueología Argentina, La Rioja. [ Links ]
23. Nielsen, A., F. Ávila y M. Vázquez. 2010. Notas Sobre la Arqueología de la Cuenca Media del Río Grande de San Juan (1000-1450 d.C.). Trabajo presentado en el XVII Congreso Nacional de Arqueología Argentina, Mendoza. [ Links ]
24. O'Connor, T. 1993. Process and Terminology in Mammal Carcass Reduction. International Journal of Osteoarchaeology 3:63-67. [ Links ]
25. Sandefur, E. 2001. Animal Husbandry and Meat Consumption. En Empire and Domestic Economy, editado por T. D'Altroy y C. Hastorf, pp. 179-202. Kluwer, New York. [ Links ]
26. Schiffer, M. B. 1975. Behavioral Chain Analysis. Activities, Organization, and the Use of Space. Fieldiana 65:103-119. [ Links ]
27. Stahl, P. 1999. Structural Density of Domesticated South American Camelid Skeletal Elements and the Archaeological Investigation of Andean Ch'arki. Journal of Archaeological Science 26:1347-1368. [ Links ]
28. Tomka, S. 1994. Quinua and Camelids on the Bolivian Altiplano: An Ethnoarchaeological Approach to Agro-pastoral Subsistence Production with an Emphasis on Agro-pastoral Transhumance. Tesis doctoral inédita, University of Texas, Austin. [ Links ]
29. Wilk, R. y R. Netting. 1984. Households: Changing Forms and Functions. En Households. Comparative and Historical Studies of The Domestic Group, editado por R. Netting, R. Wilk y E. Arnould, pp 1-28. University of California Press, Los Angeles. [ Links ]
30. Wilk, R. y W. Rathje. 1982. Household Archaeology. American Behavioral Scientist 25 (6):617-639. [ Links ]
31. Yacobaccio, H. 1994. Etnoarqueología de Pastores Surandinos: Una Herramienta Para Conocer el Registro Arqueológico. Jornadas de Arqueología e Interdisciplinas, Programa de Estudios Prehistóricos, CONICET:203-206. Buenos Aires. [ Links ]
32. Yacobaccio, H. 2007. Andean Camelid Herding in the South Andes: Ethnoarchaeological Models for Archaeozoological Research. Anthropozoologica 42 (2):143-154. [ Links ]
33. Yacobaccio, H. y M. Malmierca. 2006. The Role of the Challada in Llama Culling (Puna of Atacama, Argentina). En Kay Pacha . Cultivating Earth and Water in the Andes, editado por P. Dransart, pp. 151-156. BAR International Series 1478, Archaeopress, Oxford. [ Links ]
34. Yacobaccio, H. y C. Madero. 2001. Ethnoarchaeology of a Pastoral Settlement of the Andean Plateau: An Investigation of Archaeological Scale. En The Ethnoarchaeology of Andean South America: Contributions to Archaeological Method and Theory, editado por L. Kuznar, pp. 84-96. International Monographs in Prehistory, Ann Arbor. [ Links ]
35. Yacobaccio, H., C. Madero y M. Malmierca. 1998. Etnoarqueología de Pastores Surandinos. Grupo de Zooarqueología de Camélidos, Buenos Aires. [ Links ]

