











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink
Introduction
The genusChaetosphaeriawas established by Tulasne et Tulasne (1863) withC. innumeraBerk. & Broome ex Tul. & C. Tul. as the type species. The genus is rich in species, with 171 records in Index Fungorum (http://www.indexfungorum.org).Chaetosphaeriahad been placed in the Lasiosphaeriaceae (Barr, 1990), and it is currently in the Chaetosphaeriaceae (Réblová et al.; 1999) in the order Chaetosphaeriales (Huhndorf et al.; 2004; Maharachchikumbura et al.; 2015). As a cosmopolitan distributed genus it has been studied from both temperate and tropical regions (Hyde et al.; 1999; Fernández et Huhndorf, 2005; Fernández et al.; 2006; Réblová et al.; 1999; Perera et al.;2016). It is frequently found growing on wood or bark of different angiosperms and gymnosperms, both in terrestrial and aquatic freshwater and marine habitats (Réblová, 2004; Fernández et al.; 2006; Farr et Rossman, 2019). Both states, sexual and asexual are frequently associated (Réblová, 2000; Fernández et Huhndorf, 2005; Huhndorf et Fernández, 2005, Ariyawansa et al. 2015, Perera et al.;2016). There have been few investigations on the genusChaetosphaeriain Argentina. Peña et Arambarri (1996) describedChaetosphaeria chaetosaKohlm.; this is a marine lignicolous species, now placed inMarinokulatiE. B. G. Jones & K. L. Pang (Jones et al.; 2014).Romero (1999) describedChaetosphaeria myriocarpa(Fr.) C. Booth. andChaetosphaeria preusiiGams & Hol.-Jech. onEucalyptus viminalisLabill.
Through our work on the biodiversity of fungal species associated with the native gymnospermPodocarpus parlatorei“pino del cerro” in Argentina (Catania, 2004; Catania et Romero, 2001, 2011, 2014, 2017; Gallo et al.; 2018), a new variety and new records ofChaetosphaeriaspecies were found growing on dead wood and bark. Here we present descriptions, illustrations and comments regarding the geographic distribution for all the findings. A key to the known species ofChaetosphaeriain Argentina is also included.
Materials and Methods
The specimens were collected during mycological explorations in the forests ofP. parlatorei, in the provinces of Tucumán and Catamarca (Catania et Romero, 2010). The forests belong to the phytogeographic province of the “Yunga” of the Amazonic Dominion (Cabrera et Willink, 1980). Studied specimens are kept at mycological herbarium of Fundación Miguel Lillo (LIL); in addition, a collection kept at the herbarium PRM (Thiers, 2019) was examined.
Observations, digital imaging and measurements of microscopic structures (ascomata, asci, ascospores and asexual structures: conidiophores, conidia) were made on material mounted in distilled water, 5% KOH and 1% phloxine with either an Olympus CX31 microscope and Olympus SP-350 digital camera or a Zeiss Axioskop microscope and a Dage MTI video camera. To observe ascospores and apical rings in the asci, material was mounted in calcofluor and examined with a Zeiss Axioplan epifluorescence microscope according to Romero et Minter (1988). Drawings were made with a camera lucida (Leica MZ6).
Results and Discussion
Chaetosphaeria abietis(Höhnel) W. Gams & Hol.-Jech.; Stud. Mycol.13: 53. 1976.Zignoëlla abietisHöhnel, in Rehm, Ann. Mycol. 5: 469. 1907. Type: Austria, Dürrwien, Wiener Wald, on inner side of decayed bark, VII-1907,F. von Höhnel s/n, Rehm Ascom. exsicc. Nº 1740 (lectotype K 49555).
Fig. 1A-D
Ascomata perithecial, dark brown to black, subglobose, 150-300 × 100-250 µm, superficial, scattered to gregarious, papillate, ostiolate, shining, surrounded by sterile conidiophores. Peridium 35-45 μm thick, with cells oftextura epidermoideain surface view; in longitudinal section 2-layered; inner layer composed by elongate to flattened hyaline cells, outer layer of light brown pseudoparenchymatous cells. Asci unitunicate, cylindrical, 110-130 × 8-13 µm, 8-spored. Ascospores fusiform with rounded ends, 3-septate, straight or slightly curved, 25-32 × 3-4 µm, hyaline, each cell with a lipid droplet, smooth-walled.
Distribution and habitat:Chaetosphaeria abietiswas found in Austria and Czech Republic on wood and bark of conifers:Abies albaMill. andPicea abies(Gams et Holubová-Jechová, 1976). Réblová et Gams (1999) citedC. abietison bark ofAbies albain Czech Republic, France, Ukraine, Germany and United States of America (www.gbif.org/species).Podocarpus parlatoreiis recorded here as a new host for the species.
Specimens examined:ARGENTINA.Catamarca: Dep. Ambato, Las Juntas, in forest ofP. parlatorei, 28˚07’94”S, 65˚54’49”W, 1780 m, 26-IX-1999,Catania 1769(LIL).
Observations: Detailed descriptions ofChaetosphaeria abietiswere provided by Gams et Holubová-Jechová (1976) and Réblová et Gams (1999). Our material only differs in the size of the ascomata (ca. 180 µm diám) and of the asci (80-90(-120) × 8-10 µm) which are smaller than those described by Gams et Holubová-Jechová (1976), and in addition to the absence of a hyphal subiculum on which ascomata develop.
Chaetosphaeria abietisis reported for the first time from Argentina.
Chaetosphaeria hispidaRéblová & Seifert var.podocarpiCataniavar. nov.Type: Argentina. Tucumán: Taficillo, in forest ofP. parlatorei, 26º42’82”S, 65º19’53”W, 29-II-2000,M. del V. Catania 1811(holotype LIL). IF 555782.
Figs. 1E-H, 2A-J
Differs fromChaetosphaeria hispidavar.podocarpiby possessing longer setae (up to 47 μm) and shorter ascospores (40-49.5 μm).
Ascomata perithecial, brown to dark brown, globose to subglobose, 250-450 × 250-400 μm, superficial, scattered to gregarious, slightly papillate, ostiolate, setose. Setae dark brown, aseptate, stiff, pointed, 23-47 × 5.0 μm; abundant setae in young ascomata. Peridum 35-49.5 μm thick, with cells oftextura globosatoangularisin surface view, dark brown; 2-layered in longitudinal section, inner layer of light brown to hyaline, thin-walled flattened elongated cells, outer layer thicker, dark brown, composed of angular-polygonal cells. Paraphyses 2-3 μm wide in the base, unbranched, tapering towards the 1.5 μm wide apex, septate with intervals of 19-22 μm long. Asci unitunicate, cylindrical-clavate, 110.5-156 × 8-9(-10) μm, 8-spored, rounded at the apex, with refractive apical ring. Ascospores cylindrical-filiform, 7-septate, straight or slightly curved, with rounded ends, slightly tapering to base, (35.5-)40-49.5(-51) × 3-4 μm, hyaline, 2-seriate, smooth-walled. Conidiophores macronematous, mononematous, erect, straight or flexuous, cylindrical, unbranched, septate, smooth, brown, becoming light brown towards the apex, 221-240 μm long, 5-9 μm diam in the base, 4-6 μm diam upwards. Conidiogenous cells terminal, integrated. Conidia acrogenous, cylindrical, straight to slightly curved, apically rounded, truncated at the base, 13-17 × 5-8 μm, 3-septate, central cells brown to dark brown and distal cells pale brown to subhyaline, smooth.
Etymology: referring to the associated plant,Podocarpus.
Distribution and habitat:Chaetosphaeria hispidais known from the type collection from Thailand, on rotten wood (Réblová et Seifert, 2003). Here it is recorded on bark and wood ofP. parlatoreifrom Argentina.
Specimens examined:ARGENTINA.Tucumán: Dep. Tafí Viejo, Taficillo, in forest ofP. parlatorei, 26º42’82”S, 65º19’53”W, 1600 m, 1-III-1999,Catania 1325(LIL); idem, 24-V-1999,Catania 1661,1662(LIL); idem, 27-XI-2000,Catania 1940(LIL).THAILANDIA.Nakhon Nayok: Khao Yai National park NE of Bangkok, Bung Phai trail ca. 5 Km NW from Khao Yai forest Headquarters ona way to Pak Chong, 14º28’N 10º23’E, 750 m, MR 2220/01; on decayed wood of a branch, 6-IX-2001,Réblová, Samuels & Nasit(holotype PRM 900543).
Observations:Chaetosphaeria hispidavar.podocarpioccurs on wood and bark. Tipically, ascospores are 7-septate and scolecosporous ascospores, and the ascomal wall has atextura globosatoangulardisposition of cells in surface view.
The argentine collections agree with the morphological characteristics ofC. hispida(PRM 900543!, Réblová et Seifert, 2003) in the setose ascomata and ascospore features such as shape, number of septa and color, but differ because setae are longer (up to 47 μm vs. up to 37 μm) and ascospores shorter (40-49.5 μm long vs. 74-82 μm long), so I thus propose a new variety.
Several species ofChaetosphaeriaform anamorphs similar to those described by Réblová et al. (1999) and Seifert et al. (2011), and the variety of conidiophores and conidia when they are grownin vitrois huge (Réblová et Seifert, 2003).
Asexual structures were found around the ascomata ofC. hispidavar.podocarpi. Réblová et Gams (1999) describedC. decastyla(Cooke) Réblová & W. Gams with similar conidiophores arising from the substratum. The asexual conidiophores in the present material were considered to be related to the ascomata ofC. hispidavar.podocarpi. InC. hispida, no asexual state was observed on the natural material.
Chaetosphaeria montanaRéblová, Czech Mycol.50(3): 153. 1998. Type: Czech Republic, Southern Bohemia, Novohradske hory Mts.; virgin forest Zofinsky prales near Pivonice; on rotten wood ofFagus sylvatica,20-V-1997,M. Réblová(holotype PRM 842969. Fig. 3A-F
Ascomata perithecial, dark brown, subglobose, 150-250 × 140-200 µm, slightly immersed to superficial, scattered or in small groups, papillate; sterile conidiophores among the ascomata. Peridium 30-40 μm thick, with cells oftextura epidermoideain surface view, 2-layered in longitudinal section, inner layer of thin-walled hyaline elongate cells, outer layer of thick-walled, brown to dark brown pseudoparenchymatous cells. Paraphyses 3-4,5 µm wide, numerous, simple, filiform, septate. Asci unitunicate, cylindrical, 65-95 × 8-11 µm, 8-spored. Ascospores ellipsoid to fusiform, 1-septate to 3-septate at maturity, slightly constricted, 17-21 × 5-6,5 µm, hyaline, guttulate, smooth-walled.
Distribution and habitat:Chaetosphaeria montanawas originally found in the Czech Republic, on decayed wood ofFagus sylvaticaL. (Réblová, 1998). Up to the present, the species was known only from the type specimen, and from now, growing on bark ofP. parlatoreifrom Argentina.
Specimens examined:ARGENTINA.Catamarca: Dep. Ambato, Las Juntas, in the forest ofP. parlatorei, 28º07’94”S, 65º54’49”W, 1780 m, 16-VI-1999,Catania 1678(LIL).
Observations:Chaetosphaeria montanawas described by Réblová (1998). Our material agrees well with the original description, differing only in that both ascospores (17-21 × 5-6.5 µm vs. (18.5-) 19-25 × 5.5-7 µm) as asci (65-95 × 8-11 µm vs. 100-130 × 10.5-12.5 µm) are slightly smaller. The species is reported for the first time from Argentina, also this record constitutes the second world.
Taking into account the differences in the geographic location, habitat, host and morphological features, it would not be rare to be infront of a new species. To confirm or deny this hypothesis, more samplings and molecular and phylogenetic studies are needed.
Chaetosphaeria vermicularioides(Sacc. & Roum.) W. Gams & Hol.-Jech.; Stud. Mycol. 13: 15. 1976.Eriosphaeria vermicularioidesSacc. & Roum.; Reveu mycol. 5: 235. 1883. Type: Belgium, Malmédy, Fungi Gall. Exs. 2693 (isotype L 910.236-665).
Fig. 3G-J
Ascomata perithecial, dark brown to black, subglobose, 130-200 × 120-180 µm, superficial, gregarious, papillate, ostiole slightly conical. Peridium 20-25 μm thick,textura epidermoideain surface view, 2-layered in longitudinal section, inner layer composed of light brown elongate cells, outer layer of dark brown pseudoparenchymatous cells. Asci unitunicate, cylindrical, 45.5-61 × 3-5 µm, 8-spored, thin-walled. Ascospores ellipsoidal-fusiform, (6-)6.5-8 × 2.5-3 µm, hyaline, smooth-walled, septum in the middle, breaking apart in a rather early stage within the ascus. Conidiophores macronematous, mononematous, unbranched, septate, brown, paler towards the top, 65-130 µm × 2.5-4 µm. Conidiogenous cells phialidic, hyaline, 2.5-3 µm wide, collarette inconspicuous. Conidia ellipsoidal, hyaline, unicellular, 3.5-4(-4.5) × 2-2.5 µm.
Distribution and habitat: Belgium, Czechoslovakia, Denmark, France, Great Britain, Netherlands (Gams et Holubová-Jechová, 1976); Australia, Brazil, Germany, New Zealand, Mexico (www.gbif.org/species). Ellis et Ellis (1997) reported it on decay wood ofAlnus glutinosa(L.) Gaertn.; Fraxinus excelsiorL. andQuercussp. Gams et Holubová-Jechová (1976) mentions different substrates. Romero (1983) described the asexual state from wood and bark ofEucalyptus viminalisLabill. in Argentina. In this work, we reportChaetosphaeria vermicularioideson wood of branch ofP. parlatoreias a new host plant. The distribution area of the species is extended to the northwestern Argentina.
Specimens examined:ARGENTINA.Tucumán: Dep. Burruyacu, Sierra de Medina, ruta provincial 310, at 31 km from Villa Padre Monti, Aguas Negras, Finca Mansilla, in forest ofP. parlatorei, 26º22’06”S, 65º03’46”W, 1600 m, 22-XI-2000,Catania 1935(LIL).Buenos Aires: Pdo. de San Pedro, Gdor. Castro, on wood and bark ofEucalyptus viminalis, XI-1982,Romero 18(BAFC).
Observations: Our material agrees with the description given by Gams et Holubová-Jechová (1976), but it has slightly longer conidia (3.5-4.0(-4.5) µm vs. 2.5-3.8 µm).
Chaetosphaeria vermicularioidescan be confused withC. inaequalis(Gove) W. Gams & Hol.-Jech, but it differs mainly in ascospore size (part spores 3-4.5 µm long. vs. 2.5-3.5(-4.5) µm long).
Conidia described here are slightly larger than those in Romero (1983) (3.5-4 × 2-2.5 µm vs. 2-3 × 1.5-2 µm).
In Argentina, Arambarri et al. (1981) citedChloridium lignicola(Mangenot) W. Gams & Hol.-Jech. growing on leaf litter ofNothofagus dombeyi(Mirb.) Oerst.; in the Quetrihué Peninsula (Neuquén Province). Currently, this asexual name is considered the asexual state ofChaestosphaeria. Godeas et Arambarri (2007) described asexual structures ofChaetosphaeria lentomitaeW. Gams & Hol.-Jech.on decorticated wood ofQuercus petraea(Matt.) Liebl.;Fagus sylvaticaL.;Pinussp, from Tierra del Fuego Province.
Chaetosphaeria vermicularioidesis described for the first time for Argentina.
Key toChaetosphaeriaspecies reported in Argentina
1. Ascospores 1-septate or 1-3-septate. ......................................................................................................... 2
2. Ascospores never fragmenting into part-spores. ................................................................................... 3
3. Ascospores cylindrical with rounded ends, hyaline, 1-septate, 5-7 × 1.5-2.0 µm. Conidiophores simple, with simple phialides; conidia truncate at the base and rounded at the tip, in short chains. ....................................................................................................................................C. myriocarpa
3'. Ascospores ellipsoidal to fusiform, 1-3-septate, hyaline, guttulate, 17-21 × 5-6.5 μm. Conidiophores scattered among the sterile setae; conidiogenous cells phialides mono- or polyphialidic; conidia fusiform, falcate with pointed ends. ................................................................................C. montana
2'. Ascospores always fragmenting into part-spores. ................................................................................ 4
4. Ascospores ellipsoidal-fusiform, 1-septate, part-spores 3-4.5 × 2.5-3 µm. Conidiophores unbranched; conidiogenous cells phialidic with multiple loci apertures; conidia ellipsoidal. ......................................................................................................................................C. vermicularioides
4'. Ascospores cylindrical to ellipsoidal, 1-septate, part-spores 2.5-3.5 × 1.5-2.0 µm. Conidiophores simple; conidiogenous cells phialidic with single loci aperture; conidia ellipsoidal. .......C. preussii
1'. Ascospores 3-septate or more than 3-septate. ........................................................................................... 5
5. Ascospores fusiform with rounded ends, 3-septate, 25-32 × 3-4 μm, guttulate; ascomata glabrous. Conidiophores simple; conidiogenous cells phialidic; conidia ellipsoidal-cylindrical, base truncate. ...................................................................................................................................................C. abietis
5'. Ascospores cylindrical- fusiform, 7-septate, with rounded ends, (35.5-)40-49.5(-51) × 3-4 μm; ascomata setose. Conidiophores unbranched; conidiogenous cells terminal, integrated; conidia cylindrical. .....................................................................................................C. hispidavar.Podocarpi
Figure Captions
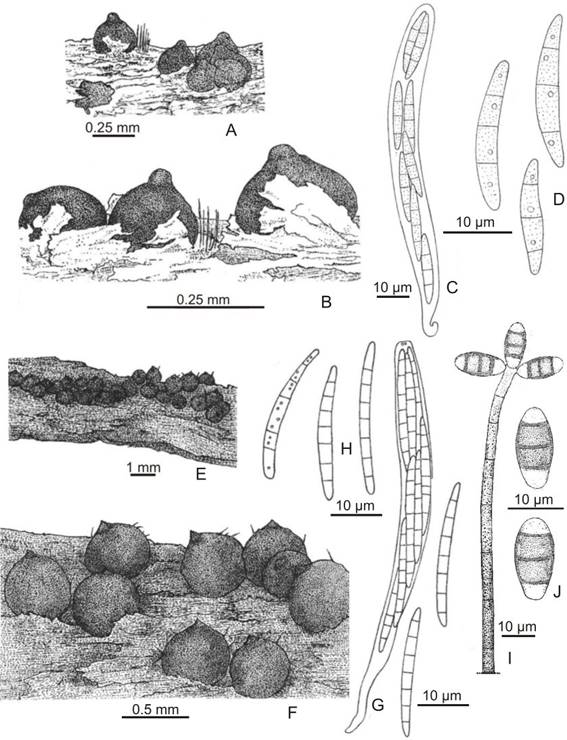
Fig. 1. Chaetosphaeria abietis. A-B: Ascomata on substrate. C: Ascus. D: Ascospores. Chaetosphaeria hispidavar.podocarpi. E-F: Ascomata on substrate. G: Ascus. H: Ascospores. I: Conidiophore, conidia. J: Conidia. (A-D,Catania 1769, LIL; E-J,Catania 1811, LIL).

Fig. 2. Chaetosphaeria hispidavar.podocarpi. A-B: Ascomata on substrate. C-D: Young ascomata with setae. E: Seta. F: Peridium, longitudinal section. G: Asci, detail of apical ring, arrow (EF). H: Ascospores (EF). I: Ascus, refractive apical ring (arrow). J: Ascospore. (A-J,Catania 1811, LIL).

Fig. 3. Chaetosphaeria montana. A-B: Ascomata on substrate. C: Ascus with young ascospore, 1-septate. D: Ascus with mature ascospore, 3-septate. E: Ascospore. F: Paraphyses. Chaetosphaeria vermicularioides. G: Ascomata, with conidiophore. H: Ascus. I: Conidiophore, conidiogenous cells phialidic. J: Conidia. (A-F,Catania 1678, LIL; G-J,Catania 1935, LIL).

