











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkCOMPLEJO MAYOR DE HISTOCOMPATIBILIDAD (MHC) EN POLLOS
En Gallus gallus, el MHC es llamado “Complejo o sistema B” por considerarse originalmente un grupo sanguíneo distinto del A, previamente descripto. Por otro lado, esta confusión se debió al hecho de que las moléculas se expresan en los eritrocitos, por ser nucleados. A través de la serología, se hallaron 28 serotipos diferentes del Complejo B en aves de postura (1 ,2). Más tarde se demostró que estas moléculas B tenían las mismas funciones que las moléculas de histocompatibilidad (3 ) , pero a pesar de ello, la nomenclatura que se utiliza para los genes del MHC en aves sigue siendo B. Por ello hay tres tipos de genes: a los genes de clase I se los denomina B-F; a los de clase II, B-L y un tercer grupo, sólo presentes en las aves de la familia de las gallináceas, de clase IV o B-G.
En los comienzos, se determinó la existencia de 6 genes homólogos a las secuencias de los genes humanos de clase I, y 5 genes o secuencias homólogas a las de clase II. Esto pudo detectarse por medio de técnicas de “chromosome walking” en una genoteca de pollos de raza Leghorn homocigotas para el serotipo B12. Por entonces se dudó si el número de copias era el mismo en los pollos de distintos serotipos o solamente se mantenía en los animales de serotipo B12. Estas 6 copias fueron llamadas del B-F I al VI (4 ) . Por medio de trabajos de hibridación con una sonda completa del gen B-F, logró descubrirse que existen solamente dos transcriptos en tejidos normales, uno con mayor tasa de expresión que el otro. De ello se planteó la existencia de sólo dos copias de los genes de clase I en el Complejo B de los pollos, los cuales son el B-F IV, llamado mayor por su tasa de expresión y el B-F I, llamado menor. Ya en la actualidad se denominan B-F I y B-F II. Se observó que los genes B-F II, III, V y VI no tienen el extremo 3’ completo, lo que impediría que se produzca una correcta expresión de estos en las células. Posteriormente se descubrió que se trataba de los genes del Sistema Rfp-Y (5 ) .
El sistema Rfp-Y codifica moléculas de clase I y II atípicas, similares a las encontradas en el MHC de mamíferos como, por ejemplo, los genes Mic. Esta afirmación se basó originalmente en el bajo polimorfismo y la baja divergencia que presentan las secuencias de los genes de dichas moléculas; como así también por el bajo nivel de expresión, la carencia de fuertes efectos en el rechazo a injertos, la pobre reacción con linfocitos y poca relación con la resistencia a enfermedades (5 ,6 ). Posteriormente, pudieron determinar que al menos una copia de los genes de clase I del sistema Rfp-Y, el YF-V, se expresa y es altamente polimórfico en el sitio de unión al antígeno (7 ) .
El Complejo B presenta la característica de ser muy compacto en cuanto a la región cromosómica que ocupa en comparación con el MHC de los mamíferos. Como ejemplo podemos señalar que todo el complejo B ocupa 250 kilobases (kb), mientras que la región del HLA (Human leukocyte antigen) ocupa aproximadamente 4 megabases (8 ) .
Las moléculas de histocompatibilidad de Clase I (B-F) son las encargadas de la presentación antigénica de los péptidos endógenos, como por ejemplo los de origen viral o tumoral, y se expresan en la superficie de casi todas las células del organismo, con excepción de los glóbulos rojos, los trofoblastos y las neuronas. Los genes de clase II (B-L) codifican las moléculas encargadas de presentar los péptidos de origen exógeno, por ejemplo: bacterias, hongos, y solo se expresan en las células presentadoras de antígenos (CPA), como macrófagos y células de Langerhans. Las moléculas de clase IV (B-G) solo se han encontrado en las aves y no pudo determinarse la existencia de una contrapartida en los mamíferos, aunque se parecen mucho a la butirofilina. Se expresan en los eritrocitos, linfocitos, células del epitelio intestinal, bursa de Fabricio y Timo, y aún no ha podido determinarse su función en forma concreta, aunque se describió una posible función coadyuvante de las moléculas B-F en la activación de los linfocitos T CD8. (9 ,10 )
ORGANIZACIÓN DE LOS GENES DEL MHC DE LOS POLLOS
El Complejo B en los pollos se halla compuesto por una serie de genes relacionados con la respuesta inmune. En el cromosoma 16 se halla la región o cluster B-F/B-L, la que contiene dos copias de los genes de Clase I y dos copias de los genes de Clase II β. Junto a los genes B-F y B-Lβ se encuentran ligados otros genes relacionados con la presentación antigénica y la respuesta inmune como, por ejemplo: una copia de los genes B-G, genes Tap 1 y 2 (encargados de la carga del antígeno en el surco de las moléculas de Clase I), gen C4 del complemento, etc. Un mapa simplificado se muestra en las Figuras 1 y 2 .
MARCADORES MOLECULARES DEL MHC
Las determinaciones serológicas presentan una buena correlación con la caracterización molecular de los genotipos del MHC. El análisis a nivel molecular de los genes que conforman el MHC presenta una ventaja adicional, ya que por medio de estos tipos de análisis es posible detectar mejor la variabilidad de los genes de clase I y II, que pueda pasar inadvertida en las determinaciones serológicas. La serotipificación se ha desarrollado principalmente en líneas consanguíneas de aves de postura (Leghorn) y los antisueros no han sido muy efectivos a la hora de caracterizar líneas de aves para carne, por tal motivo la caracterización de los genes del MHC es una herramienta de fundamental importancia en el estudio de los pollos para carne
El microsatélite LEI0258 se halla ligado aproximadamente a 50.000 pares de bases (pb) de los genes B-F en el cromosoma 16 de las aves. Esta distancia equivale a 0,5 centimorgan (cM), lo que indica que es un buen marcador del complejo B.Se ha encontrado una asociación entre los patrones de herencia de este microsatélite con los 28 serotipos descriptos (2 ) . El microsatélite tiene pocos alelos y los serotipos homocigotas tienen una sola banda de amplificación de este microsatélite, mientras que los heterocigotas presentan dos. Según el tamaño de las bandas y del número de repeticiones del microsatélite puede distinguirse a los distintos serotipos y los genotipos de los animales analizados (13 ,14 ). Para nuestra población en estudio esto es de gran utilidad debido a que los anticuerpos generados en aves de postura no dan reacciones específicas en pollos para carne. El microsatélite LEI0258 es una región repetitiva de la siguiente secuencia de 12 o 13 pares de bases: (CCTTCTTTCTTTn) (14 ). Esta secuencia se repite un número variable de veces (n) y está flanqueada por regiones conservadas, lo que permite seleccionar oligonucleótidos apropiados para la amplificación de dicho marcador por la técnica de reacción en cadena de la polimerasa (PCR).
Cada animal hereda una copia del microsatélite de cada uno de sus progenitores, lo que permite distinguir la herencia de los patrones de dicho microsatélite y por ende el haplotipo al cual está ligado.
RELEVANCIA DEL ESTUDIO DE LOS GENES DE CLASE I DEL MHC EN POLLOS
El MHC ha mostrado tener una gran importancia en la determinación de la resistencia/susceptibilidad en aves a distintos patógenos como: bacterias, parásitos y virus (15 ,16 ).
Se ha encontrado una asociación entre determinados serotipos del MHC y la resistencia o susceptibilidad a la Enfermedad de Marek (16 ) . Ésta es una enfermedad linfoproliferativa de las aves causada por un herpesvirus, el virus de la enfermedad de Marek (MDV: del inglés Marek Disease Virus). Esta enfermedad se caracteriza por desmielinización del sistema nervioso periférico y linfoma de células T. Se ha demostrado que los pollos que presentan el serotipo B21 por medio de detección serológica, son resistentes a la enfermedad de Marek, mientras que los individuos con serotipo B19 o B13 son susceptibles a la enfermedad (17 ) . La mortalidad producida por la infección con el MDV es baja, sin embargo, presenta una alta morbilidad y las lesiones que se producen en los pollos afectados son una de las principales causas de la reducción en la calidad y en los niveles de producción avícola. Esta enfermedad se controla por medio de la vacunación, siendo ésta más efectiva, en animales genéticamente resistentes al virus (18 )
Se reconocieron tres mecanismos principales de la resistencia genética a Marek: la resistencia asociada al MHC, la resistencia asociada a genes MHC atípicos (Rfp-Y) y la resistencia dada por la variación en la susceptibilidad de las células T a la transformación por parte del virus (15 ,19 ).
Dada la importancia económica que representa la obtención de animales genéticamente resistentes a distintas enfermedades, se han seleccionado líneas de pollos resistentes y susceptibles a la enfermedad de Marek (20 ) . En este caso particular, los animales serológicamente conocidos como B21, poseen una tasa de supervivencia del 85 % frente a la descarga con el virus Esta capacidad que presentarían algunos serotipos del MHC en otorgar resistencia a determinados patógenos estaría explicada por la teoría del Mínimo Esencial planteada por James Kaufman (8 ) . Esta teoría plantea que, al ser las moléculas de los pollos tan poco variables en comparación con las de los mamíferos, existirían en la naturaleza alelos y copias de los genes de histocompatibilidad que son “mejores” en la presentación antigénica de los péptidos virales. Esta capacidad para presentar péptidos endógenos de un modo más eficiente por parte de las moléculas de clase I (B-F) daría a esa variante o serotipo una mayor capacidad del sistema inmune para responder, y en consecuencia, una resistencia de los animales ante las patologías. Por otro lado, en caso de que la variante de los genes no sea una “buena presentadora antigénica”, la respuesta inmune será menor y los animales serán más susceptibles al patógeno.
En cuanto a los pollos para carne, se han realizado pocos estudios al respecto. El grupo que comenzó con estos estudios para tratar de lograr serotipificación de líneas de carne fue el de Sandy Ewald en Auburn, EE. UU. En una línea comercial de pollos para carne se determinaron 3 serotipos del MHC. A esta línea se la denominó línea C, nombre fantasía con el fin de preservar la información genética de dichas líneas (12 ). El mismo grupo de trabajo logró determinar secuencias parciales de la región α1 y α2 de los genes B-F de las líneas para carne denominadas A, B y C. En ellas se encontraron, como era de esperar secuencias pertenecientes a haplotipos únicos en parrilleros como el C2v, A12, C7 entre otros (21 ,22 ).
De los tres genes mayores (moléculas que presentan antígenos) que conforman el MHC de las aves (B-F, B-L y B-G), se ha encontrado que existe una correlación entre los distintos genotipos de los genes B-F y la existencia o no, de susceptibilidad a Enfermedad de Marek. Se observó que los animales recombinantes (B-G21, B-F19) fueron igualmente susceptibles al virus de la Enfermedad de Marek que los pollos de serotipo B19. Los genes asociados al Complejo B e involucrados con la resistencia a la Enfermedad de Marek fueron localizados en la región B-F/B-L del cromosoma 16 (23 ) . Sin embargo, otros trabajos describen diferencias en la tasa de mortalidad en animales de idéntico genotipo para los genes B-F y que presentaban diferencias en los genes B-G (24 )
Del mismo modo que con la serotipificación, la mayoría de los trabajos que se han hecho estudiando la relación existente entre el MHC y la resistencia a Enfermedad de Marek, fue llevada a cabo en aves de postura. La caracterización de los genes de Clase I, a través de técnicas de biología molecular, es de importancia debido al hecho de que se trata de una población no caracterizada serológicamente. Esta caracterización se realizó por medio de las técnicas de: Polimorfismo de Longitud de los Fragmentos de Restricción (RFLP del inglés: Restriction Fragment Length Polymorphism) y Polimorfismo Conformacional de Cadena Simple (SSCP del inglés: Single Strand Conformation Polymorphism) en la línea AH de Pollos Campero-INTA (25 ) así como en otras poblaciones, demostrando su utilidad (26 ).
La determinación de secuencias de los genes de clase I por medio de secuenciación automática es una técnica muy precisa, pero muy costosa. Estas secuencias se realizan en animales de líneas homocigotas, lo que ha generado una gran cantidad de reportes existentes en el GenBank de los transcriptos de genes B-F de la mayoría de los serotipos conocidos.
Recientemente en 2016, Fulton et al. (27 ) desarrollaron un panel de análisis de la variación de 90 polimorfismos de un solo nucleótido (SNPs) abarcando la región más importante del MHC hasta llegar a la región de los genes B-G, sin incluirlos. Esta región contiene a los genes de clase I y clase II y sus genes intermedios, alrededor de 16 de ellos en su mayoría involucrados en la respuesta inmune. De esta forma, el análisis de la variabilidad en distintas razas y líneas se hace mucho más completo y preciso.
TÉCNICAS UTILIZADAS PARA LA DETECCIÓN DE GENOTIPOS Y HAPLOTIPOS DEL MHC
Se han utilizado varias técnicas basadas en ADN para examinar la diversidad MHC-B en pollos. La técnica de Southern blot que utiliza sondas específicas de genes más importantes del MHC-B ha revelado gran diversidad MHC-B en muchas razas de pollos. (25 , 28 –3 0 ) La técnica de SSCP (Single-Strand Conformation Polymorphism) se ha utilizado para examinar pequeñas regiones de genes específicos (2 6) . La secuenciación directa de genes específicos también se ha utilizado para identificar los haplotipos (12 ,31 ,32 ) .Estas técnicas fueron particularmente útiles para examinar la variación de MHC en pollos de diferentes razas. Sin embargo, todas estas técnicas no son adecuadas para el análisis de un gran nùmero de animales y además son laboriosas y costosas.
El microsatélite LEI0258 es una región STR (repeticiones cortas en tándem) situada dentro del MHC-B: LEI0258 contiene repeticiones en tándem de 12 y 13 pb más varias deleciones en la región flanqueante que dan lugar a las diferencias de tamaño que determinan los distintos alelos (13 ,14 ).
Debido al gran tamaño de las dos repeticiones en tándem, la variación de tamaño alélico es grande. Los alelos pueden distinguirse fácilmente utilizando electroforesis para separar los productos de PCR en geles de agarosa. Casualmente, los tamaños de los alelos del marcador coinciden con los serotipos definidos previamente, por ejemplo, el haplotipo B21 tiene un alelo de 357 pb permitiendo inferir el serotipo. Chazara et al., 2013 (33 ) detectaron 79 alelos LEI0258 en un estudio realizado a 80 poblaciones de pollos de varios países. El marcador LEI0258 se ha utilizado para identificar la variación de MHC-B en las líneas Leghorn, de postura de huevos así como en biotipos de pollos originarios de múltiples países (14 ), incluyendo África, Irán, Corea, India y en Argentina en líneas sintéticas INTA (27 ,3 4 y 39 ).
Aun cuando sigue utilizándose el marcador LEI0258 por su simplicidad y bajo costo, hay limitaciones para la detección de haplotipos del MHC-B con el mismo. Diferentes haplotipos MHC-B pueden tener el mismo tamaño de alelo LEI0258, de modo que no siempre es preciso. Hay algunos serotipos que dan dos tamaños de bandas con el marcador. Dado que los microsatélites poseen una alta tasa mutacional, la mutación en el marcador puede dar lugar a diferentes tamaños de alelos LEI0258 que se encuentran dentro de los mismos haplotipos MHCB (14 ,27 ). Además, pueden producirse diferencias en las estimaciones detamaño de alelo realizadas en diferentes laboratorios y confundir laclaridad de las asignaciones de tipo. Dado que LEI0258 es un marcadorúnico, solo se evalúa una posición dentro del MHC.
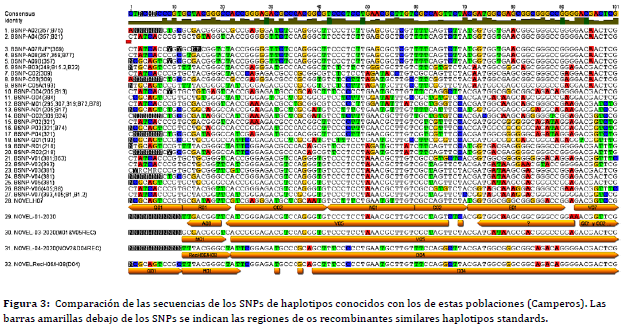
Más recientemente se ha desarrollado una nueva metodología de detección de SNPs para analizar la región del MHC-B. Un panel de 90 SNPs que abarca 210.000 pb de la región de pollo MHC-B fue desarrollado inicialmente por Chazara et al. (40 ) y perfeccionado luego por Fulton et al. (27 ). Este panel fue posteriormente modificado y ampliado para incluir un gen BG adicional en uno de los extremos y validado en varias pruebas de genotipado con múltiples líneas de pollo, razas de locales y pollos de aves silvestres de las que se supone que deriva el pollo doméstico (Red Jungle fowl) (27 ,41 ,42 ). El panel SNP, que incluye al marcador VNTR LEI0258, ha hecho que sea fácil distinguir los haplotipos, identificar nuevos recombinantes y definir la variación de MHC en pollos de múltiples fuentes. Este panel de SNPs demostró que es especialmenteútil para analizar la variabilidad de MHC y establecer haplotipos denominados BSNPs (38 ,3 9,42 ,43 ).Cada animal tiene dos haplotipos, cada uno da como resultado una combinación heredada de cualquiera de los padres. Por lo tanto, en nuestros estudios publicados con pollos Campero-INTA (38 y39 ) tratando de utilizar ambas metodologías: un primer escaneo con LEI0258 y posteriormente, el panel SNP descrito por Fulton et al., 2016, para identificar posibles recombinantes o nuevos haplotipos (27 ). La figura 3 muestra un ejemplo de cómo se comparan los SNPs de todas las posiciones indicadas en la figura 2 y cómo se evalúan los nuevos haplotipos incluyendo los posibles recombinantes (Figura 3).
LÍNEAS SINTÉTICAS DE POLLOS PARA CARNE INTA
La cría de pollos para carne necesita una gran inversión de capital para formar parte del sistema de integración que rige el mercado del pollo en galpones industriales y dio lugar a que muchos pequeños y medianos productores fueran marginados de él, a pesar de tener experiencias y las instalaciones mínimas para la cría. La búsqueda de productores naturales o ecológicos está creciendo en todo el mundo como resultado de la convicción de que el desarrollo económico debe ser ineludiblemente sostenible, es decir, satisfacer las necesidades de la generación actual sin comprometer generaciones futuras, y también por una revaloración de la “comida sana y natural”, que está vinculada al surgimiento de la conciencia ecológica. Esta tendencia y la buena aceptación de los productos ecológicos en las góndolas de supermercado, augura un buen futuro para este tipo de producción. La línea de pollos sintética “Campero” fue desarrollada por INTA (Instituto Nacional de Tecnología Agropecuaria) en la década de los 80’s en la estación experimental ubicada en Pergamino, Buenos Aires, Argentina por el Ing. Manuel Bonino, más adelante junto a la Dra. Zulma E Canet, quien dirige actualmente el programa PROHUERTA nacional. El programa PROHUERTA es un programa de políticas públicas que promueve las prácticas productivas agroecológicas para el autoabastecimiento, la educación alimentaria, la promoción de ferias y mercados alternativos con una mirada inclusiva de las familias productoras. El Campero deriva de cruces de razas estándar para proporcionar aves de corral de crecimiento más lento que funcionan bien con terminación a pasto, sin un insumo significativo de alimento y que estuviera bien adaptado a las condiciones locales en las condiciones de cría traspatio de Argentina (11 ).
También INTA, a través de la sección avícola de la EEA INTA Pergamino, ha desarrollado y promovido el desarrollo de un protocolo para la producción del Campero, basado en la normativa francesa para la producción del pollo “Label Rouge” que en ese país ocupa hasta un 30 % del mercado de la carne aviar. Este tipo de animal representa otra oportunidad para generar un producto diferenciado por un sistema de producción menos intensivo. Esto permitirá la conversión de los pequeños y medianos productores a un sistema de producción más natural para producir pollos traspatio, ya que no requiere una sofisticada tecnología de control ambiental, sino condiciones de reproducción semi-intensivas, con menos inversión de capital.
ASOCIACIONES PREVIAS CON LA RESPUESTA INMUNE O LOS RASGOS DE PRODUCCIÓN DE LOS HAPLOTIPOS ENCONTRADOS EN LÍNEAS DE CAMPERO INTA
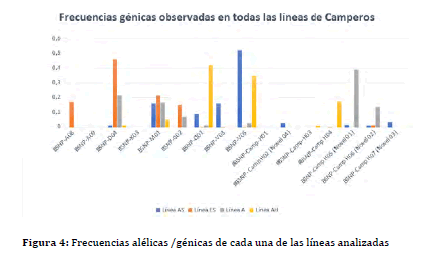
Tanto las frecuencias de cada haplotipo (alélicas) como los haplotipos hallados en estas líneas sintéticas están resumidas en la Figura 4 . En las poblaciones analizadas por esta metodología y que fueron descriptas previamente, (38 ,39 ) se hallaron varios haplotipos, algunos ya descriptos como el BSNP-D04 (conocido previamente como serotipo B13) y que tiene un patrón de tamaño del LEI0258 de 205 pares de bases (pb), se describió como uno de los que posee pobre resistencia a Marek y otras enfermedades y con resistencia moderada a enfermedad infecciosa bursal (IBD) (44 ).
En la bibliografía el alelo de LEI0258 de 205 bp se asoció significativamente con las respuestas elevadas de anticuerpos primarios contra la vacuna contra el NDV (Newcastle disease virus), (45 ) pero la línea B13 tuvo una mortalidad del 60 % y del 100 % en dos ensayos separados (46 , 47 ). Sin embargo, los resultados de estos ensayos deben mejorarse porque en otras publicaciones se mostró un crecimiento tumoral diferencial con un 94 % de regresión tumoral en las aves heterocigotas B6/B13 en comparación con la regresión tumoral del 4 % en aves homocigotas para B13/B13 en los casos de tumores inducidos por el virus del sarcoma de Rous (RSV) según Schierman et al., 1977 citado por Miller et al., 2016 (48 ) . Curiosamente, el haplotipo B13 confiere más resistencia a la celulitis causada por las infecciones por Escherichia coli en comparación con B2. A pesar de estos resultados de susceptibilidad a algunas enfermedades, el serotipo B13 se asoció positivamente con el peso corporal(49 ). Se lo describe también como BF2*0401 del haplotipo B4 (50 ) .
La banda de amplificación de 381 bp de LEI0258, que se encuentra en estas líneas sintéticas INTA (BSNP-V03 y V05, NOVEL 01, 02 y 03) (39) también se encontró en la línea AH (38 ). La región del exón 2 B-F que fue secuenciada en 2002 (Iglesias, datos no publicados) al amplificar el LEI0258 da una banda de amplificación de 381 pb. Esta secuencia resultó ser idéntica a “C2v” reportada por Livant y Ewald (22) en líneas de pollos de carne (51 ). Este alelo se encuentra positivamente asociado al peso corporal (52 ). El alelo de 307 pb del LEI0258 (BSNP-O02 B24) se encontró positivamente correlacionado con los rasgos de peso corporal en algunas poblaciones de pollos (4 5) mientras que el alelo de 309 pb pareciera poseer una asociación negativa con la supervivencia y la producción de huevos hasta las 72 semanas de edad (53 ).
Como se ha resaltado varios de los haplotipos hallados están correlacionados con mayor peso corporal sugiriendo que una selección por el peso ha sido realizada a lo largo de los años. Sin embargo, aún no se han realizado selección por resistencia genética a enfermedades que es uno de los objetivos a los que apuntan estos estudios, aprovechando los haplotipos hallados o incorporando nuevos a las líneas parentales. Deberían realizarse también estudios de asociación con la respuesta inmunitaria y rasgos productivos, para continuar en la caracterización de las líneas que originan el pollo Campero INTA como en otras poblaciones, especialmente aquellas que realizan programas de cría y mejora genética entre estas líneas. Por ejemplo, podría analizarse la respuesta inmune después de la vacunación, la asociación entre haplotipos y prevalencia de coccidiosis aviar, u otros parásitos, o, probar infecciones virales o bacterianas, ya sea en animales vivos como en células embrionarias de pollo.


Agradecimientos
A la Universidad Nacional de Río Negro por el aporte económico en los proyectos de investigación 40-A-498 y 40 A 485, sin los cuales no hubiera sido posible las publicaciones que han contribuido a esta revisión. A la Dra. Janet Fulton por su constante apoyo a estos trabajos y su generosidad por la colaboración con nuestro grupo. A Jesús Arengo y María Pía Beker por sus aportes y la lectura detallada de este escrito A todos los alumnos y becarios de la Universidad Nacional de Río Negro que han colaborado con el procesamiento de muestras.
