











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
La calidad de sitio, en bosques, es la capacidad productiva de un área terrestre para sustentar el crecimiento del mismo (González et al., 2004; Aguirre et al., 2022), presentando en algunos casos bosques exuberantes mientras que, en otros bosques pobres (Adeyemi, 2016); asimismo, es el potencial de un determinado sitio que aporta los recursos para la producción de madera u otro producto para una especie o grupo de especies (Clutter et al., 1983; Chagas y Garcia, 2017). Estás diferencias pueden ser producto de factores edáficos, topográficos y climáticos, viéndose reflejado en las especies como también en el desarrollo estructural del bosque (Adeyemi, 2016). La calidad de sitio se puede determinar por métodos directos (fitocéntricos) e indi- rectos (geocéntricos) (Prodan et al., 1997; Weiskittel et al., 2011). El método directo requiere información de crecimiento del rodal, cuyos datos son obtenidos mediante inventarios forestales continuos (Molina et al., 2019).
El método indirecto se basa en la medición de parámetros del sitio que afectan el crecimiento del bosque como el suelo, clima, vegetación y/o fisiografía (Gadow et al., 2007; Mora et al., 2015). Pocas investigaciones abordan la caracterización de los sitios forestales (García et al. 2017), la mayoría se enfoca en los métodos directos (Zhu et al., 2019; Sharma, 2021; Guerra et al., 2021).
Los estudios de calidad de sitio, son importantes porque permiten correlacionar las características del sitio con el crecimiento del bosque (Bontemps y Bouriaud, 2014); esencialmente, expresan y cuantifican los múltiples efectos de las variables o factores ambientales sobre el crecimiento de la altura máxima o dominante de los árboles (González et al., 2013; Bontemps y Bouriaud 2014; Menéndez et al., 2015). La ausencia o presencia de algunas características ambientales pueden generar limi- taciones o impulsar el crecimiento de una especie (Kiviste, 2002; Gadow et al., 2007; Mora et al., 2015); en ese sentido, contar con el conocimiento del sitio resulta fun- damental; puesto que permite conocer los requerimientos ambientales de la especie, para su crecimiento y desarrollo, a partir de las cuales se pueden seleccionar áreas propicias para instalar plantaciones forestales (González et al., 2013; Liu et al., 2021).
El género Cinchona L. (Rubiaceae-Cinchoneae), está conformado por 23 especies de árboles o arbustos (Ulloa et al., 2017; Aymard, 2019), reportándose 19 para Perú (Suáres, 2020), de las cuales 11 especies se encuentran en el Norte del país (Caja- marca y Piura), representando un 48 % del total de especies del género (Huamán et al., 2019). Cinchona, conocida comúnmente como “árbol de la quina” o “cascarilla”, tiene un gran valor cívico plasmado en el escudo nacional, que representa la riqueza vegetal del Perú (Cóndor et al., 2019). Los bosques montanos, hábitat de Cinchona, son afectados por la quema y el crecimiento urbano, construcción de vías, expansión agrícola y ganadera (Rasal et al., 2012; Aymard, 2019; Huamán et al., 2019).
En Perú, se ha promulgado la R.M. N° 0258-99-AG que prohíbe en todo el país la extracción de especímenes del género Cinchona en bosques naturales y el D.S. N° 043-2006-AG que declara en estado vulnerable a C. calisaya Wedd., pero es mínimo su cumplimento a pesar de los esfuerzos de las autoridades para la protección de las especies de este género. A pesar de la presión a la que están sometidas las especies de Cinchona (García et al., 2022), es poca la información sobre aspectos ecológicos, entre ellas C. fruticosa L. Andersson, C. glandulifera (Ruiz) Ruiz & Pav., C. krauseana L. Andersson, C. pyrifolia L. Andersson y C. scrobiculata Bonpl., están clasificadas como datos insuficientes (DD) y C. micrantha Ruiz & Pav. como preocupación menor (LC) (Pino y Taylor, 2006; IUCN, 2022). Teniendo en cuenta que el hábitat del género está siendo seriamente afectado por acciones antrópicas, tal como se evidencia en el bos- que La Palma, Chota, y la falta de investigaciones en sus requerimientos (Huamán et al., 2019), específicamente sobre la calidad de sitio, la presente investigación tuvo como objetivo evaluar y estimar la calidad de sitio de Cinchona pubescens Vahl. en el bosque montano La Palma, provincia de Chota, Cajamarca (Perú), lo que permitirá generar planes de poblamiento y recuperación.
MATERIALES Y MÉTODOS
Ubicación
El departamento de Cajamarca se ubica al Norte de Perú tiene una extensión terri- torial de 32.967,20 km2, por el Norte limita con Ecuador, por el Este con el depar- tamento de Amazonas, Sur con la Libertad y al Oeste con las regiones de Lamba- yeque y Piura, está conformado por 13 provincias, la provincia de Chota se ubica al Norte de Cajamarca y al Este de Lambayeque tiene una extensión de 3.760,06 km2. El área de estudio La Palma se localiza en el distrito y provincia de Chota, en la región Cajamarca, entre las coordenadas 6° 29’ 27,84” S y 78° 37’ 59,80” W (Fig. 1). Pertenece a la zona vida de Bosque Húmedo Montano Bajo Tropical (bh-MBT), con pendientes de ondulado suaves a onduladas, los suelos se caracterizan por ser profundos a superficiales, franco arenosos, con drenaje moderado y erosión ligera a muy ligera. El área de estudio cuenta con una extensión de 25 ha, se ubica a 40 minutos de la ciudad de Chota, de la cual se accede a través de la carretera hacia el distrito de Chalamarca.

Fig 1. Localización del área de estudio: A) Ubicación de la región Cajamarca en el territorio peruano.B) Detalle de la ubicación de la provincia de Chota en la región Cajamarca. C) Ubicación del área de estudio en el bosque montano La Palma.
Fig. 1. Location of the study area: A) Location of the Cajamarca region in Peru. B) Detail of the location of the province of Chota in the Cajamarca region. C) Location of the study area in La Palma montane forest.
Instalación de parcelas y colecta de datos
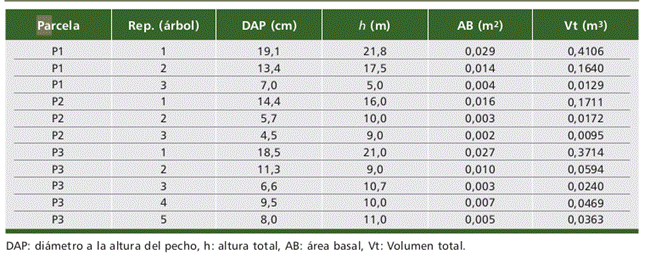
Las parcelas fueron instaladas mediante el diseño de muestreo no probabilístico por conveniencia (Cruzado et al., 2019), la instalación de parcelas garantizó la pre- sencia de individuos de C. pubescens. se instalaron a 50 m entre sí debido a la poca presencia de la especie y a la intervención antrópica que presenta el área de estudio. Se establecieron tres parcelas circulares de 500 m2 (r = 12,62 m). De cada árbol, se registró la circunferencia a la altura del pecho (CAP) medido a 1,30 m sobre el nivel del suelo con cinta métrica y se estimó la altura de árbol (h) con hipsómetro Suunto Pm-5/360. En total, 11 árboles fueron medidos. El diámetro a la altura del pecho (DAP), Área Basal (AB) y Volumen total (Vt) fueron determinados mediante las siguientes fórmulas (Soares et al., 2011):

Determinación de clases de sitio
Teniendo en cuenta que la calidad de sitio es igual al promedio de la altura de los 100 árboles dominantes por hectárea (West, 2015), en cada parcela de 500 m2 (0,5 ha) se tendrían que haber evaluado cinco árboles; sin embargo, debido a las características de la especie y la dinámica del bosque, se consideraron todos los individuos de cada parcela para la evaluación.
El análisis estadístico fue realizado en el programa R Core (Team, 2022). La existencia de clases de sitio para C. pubescens, se determinó mediante el Análisis de Varianza (ANOVA) de las alturas totales (h) en las parcelas evaluadas (Bravo et al., 2011; Soto, 2019; Villena et al., 2022). El ANOVA fue realizado a un 95% de probabilidad (p < 0,05) a través de la función Aov de R (Chambers et al., 1992). La normalidad y homogeneidad de varianza se determinó usando las pruebas de Shapiro Wilk y Barlett (p < 0,05), mediante la función shapiro.test (Royston, 1982) y bartlett.test (Bartlett, 1937) respectivamente. Asimismo, con el fin de reforzar los datos obtenidos se realizó el ANOVA del DAP y, AB; en el caso del Vt que no presentó normalidad, se usó la prueba no paramétrica de Kruskal-Wallis (p < 0,05) con la función kruskal. test (Hollander y Wolfe, 1973)
Datos edafoclimáticos
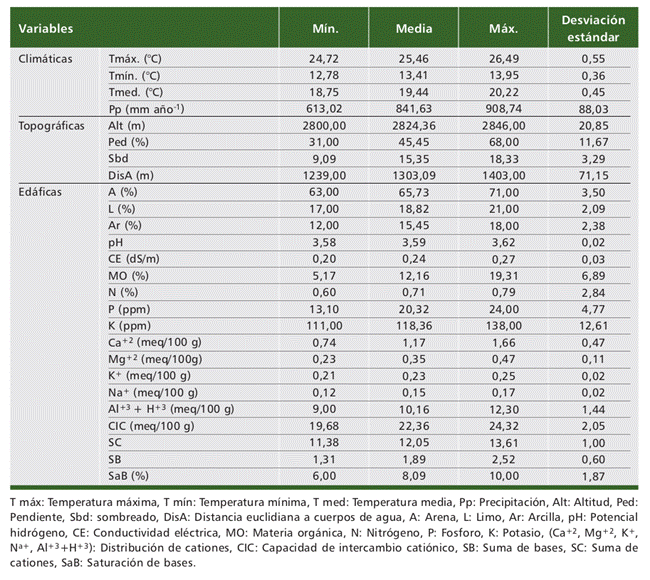
Considerando que la temperatura y la precipitación son los elementos climáticos que ejercen alta influencia sobre el crecimiento de ecosistemas boscosos (Montero, 1999). Las variables climáticas para el área de estudio fueron temperatura mínima (Tmín.), temperatura media (Tmed.), temperatura máxima (Tmáx.); obtenidas de la base de datos The POWER Project (NASA, 2021) disponibles en el sitio (https:// power.larc.nasa.gov/). La precipitación (Pp) fue obtenida de la base de datos Giovanni (https://giovanni.gsfc.nasa.gov/giovanni/). Se analizaron los datos climáticos desde el 2009 hasta 2019 (Tabla 2).
Asimismo, para cada parcela se determinó la altitud mediante un GPS y la pendiente (Ped) con eclímetro. El sombreado (Sbd) fue obtenido a partir de un Modelo Digital de Elevación (MDE) de 12,5 m de resolución espacial y la distancia euclidiana a cuerpos de agua (DisA) fue determinada a partir del shapefile de la red hídrica (GORECAJ, 2022), utilizando software de sistemas de información geográfica (SIG), Tabla 2.
Para determinar las variables edáficas, se abrieron calicatas, una por parcela, describiendo los horizontes por observación directa. Se extrajo una muestra de suelo de 1 kg por calicata, hasta una profundidad de 40 cm, debido a que un gran porcentaje de plantas cuentan con mayor densidad radicular en los primeros centímetros de la superficie (Mendoza y Espinoza, 2017), y que son estas las responsables de la absorción de nutrientes y agua (Herrera y Alvarado, 1998). Las muestras fueron colectadas y etiquetadas en bolsas tipo ziploc.
El análisis fisicoquímico del suelo se realizó en el Laboratorio de Análisis de Suelos, Plantas, Aguas y Fertilizantes de la Universidad Nacional Agraria La Moli- na, cuyos resultados permitieron conocer la textura (Arena - A, Limo - L y Arcilla - Ar), el pH, conductividad eléctrica (CE), materia orgánica (MO), nitrógeno (N), fósforo (P), potasio (K), calcio (Ca+2), magnesio (Mg+2), sodio (Na+), aluminio (Al+3+ H+), capacidad de intercambio catiónico (CIC), suma de cationes (SC), suma de bases (SB), saturación de bases (SaB), además de calcáreo total y acidez de cambio (Tabla 2).
Para determinar las variables edafoclimáticas que influyen en el crecimiento de la altura de los árboles (h), se realizó el análisis de correlación lineal de Pearson (p ≤ 0,05), efectuándose con la función cor de R (Becker et al., 1988). Finalmente, con el fin de tener un análisis global de las variables del sitio en el que se desarrolló la especie, se efectuó un análisis multivariado mediante Análisis de Componentes Principales (ACP) del paquete FactoMineR y factoextra (Lê et al., 2008; Kassambara, 2019) de R, se utilizó el método Biplot que permitió una representación gráfica de los individuos de cada parcela y las variables edafoclimáticas evaluadas (Pardo, 2020).
Caracterización de la calidad de sitio de C. pubescens
Para caracterizar la calidad de sitio de C. pubescens, se aplicó el método indirecto, considerando los factores climáticos, edáficos y topográficos, debido al desconoci- miento de la edad de los árboles (Bravo et al., 2011; Mora et al., 2015; Hargreaves et al., 2020). La caracterización fue realizada teniendo como base la capacidad de uso mayor (CUM) según los criterios establecidos en el D.S. Nº 005-2022-MIDAGRI (MIDAGRI, 2022).
Tabla 1 Características dasométricas de los individuos de C. pubescens, observados en las parcelas de medición.Table 1. Dasometric characteristics of C. pubescens individuals observed in the measurement plots.
RESULTADOS Y DISCUSIÓN
Determinación de clases de sitio
La calidad del sitio se puede estimar conociendo las propiedades físicas, químicas y biológicas del suelo (Subedi y Fox, 2016; Sotomayor, 2023); pero generalmente se recurre a la expresión cuantitativa de la calidad de sitio, siendo el más común la altura de los árboles de mayor crecimiento (Prodan et al., 1997) o altura dominante, que fue utilizada por primera vez como magnitud de la calidad de sitio por Baur (Alvarado et al., 2013). En la presente investigación se determinó que en la zona de estudio no existe diferencia significativa entre la altura media de los individuos de las parcelas (p < 0,5761), infiriendo que las alturas totales de los árboles de las tres parcelas evaluadas son estadísticamente similares (Tabla 3); con un coeficiente de variación de 45,91%. Esto indica que en el área evaluada existe una sola clase de sitio
(I) para C. pubescens, estos resultados son similares a los obtenidos en el bosque de protección Pagaibamba para Cinchona officinalis L. (Villar et al., 2018) y, en Tara spino- sa (Feuillée ex Molina) Britton & Rose en bosques naturales de Cajamarca (Villena et al., 2022). De igual forma para el DAP, AB y Vt no existió diferencias significativas entre parcelas, reforzando la existencia de sólo una clase de sitio. Estos resultados no diferenciales entre las características de los individuos de C. pubescens, probablemente se debe a que los bosques montanos son peculiares en su topografía, con variados periodos de precipitación, humedad relativa, y exposición hacia el Oeste (océano Pacífico), presentando influencia de la selva alta, situada al Este del río Marañón (Henderson et al., 1991; León et al., 2006). Factores que han permitido un proceso adaptativo de todas las especies, incluida la C. pubescens, a crecer y desarrollar en estos ecosistemas, razón por la cual no se aprecian diferencias significativas entre los individuos evaluados.
Donde, h: altura total de árbol, DAP: diámetro a la altura del pecho, AB: área basal, Vt: volumen total.
Tabla 3 Comparación de medias de h y otras variables dasométricas. Table 3. Comparison of means of h and other dasometric variables.

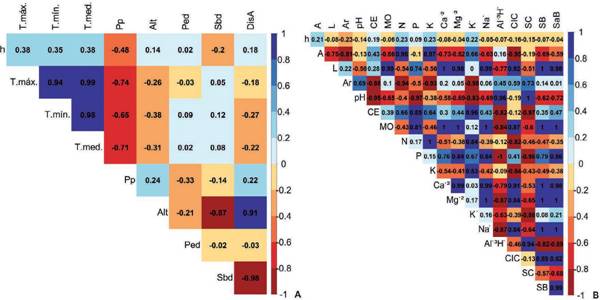
Fig. 2 Correlación entre las variables en estudio: A) Variables climáticas y topográficas vs. la altura total. B) Variables edáficas con respectos vs altura total.Fig. 2.Correlation between the studied variables: A) Climatic and topographic variables vs. total height. B) Edaphic variables with respect to total height.
Influencia de las variables edafoclimáticas y topográficas sobre el crecimiento en altura total de C. pubescens
En el análisis de correlación, no se evidenciaron correlaciones lineales entre los fac- tores edafoclimáticos y topográficos con respecto a h de C. pubescens. Sin embargo, se observó que h presentó una asociación moderada con las variables Tmáx (0,38), Tmín (0,35), Tmedia (0,38) y Pp (-0,48) (Fig. 2).
La variabilidad meteorológica interanual es un factor que influye en los patrones de actividad fisiológica de las plantas (Murphy et al., 2020), mientras que el incre- mento de algunos parámetros que afectan el cambio climático ha permitido conocer que los factores climáticos ejercen influencia vital en el crecimiento de los árboles (Duan et al., 2022; Campos, 2023); asimismo, las variables topográficas y climáticas ejercen influencia sobre la productividad de un sitio, donde la temperatura juega un papel primordial en la generación de microambientes, acelerando la descomposición de MO, que provoca la liberación de nutrientes (Martínez et al., 2013). A su vez, la temperatura también se correlaciona con la productividad de un sitio (Farrelly et al., 2009), por lo que podría tener influencia en la distribución espacial de las especies de vegetación natural (Lozada y Sentelhes, 2008). Existe evidencia que la tempera- tura es un indicador de la disponibilidad de nutrientes, al aumentar, se incrementa la absorción activa de los elementos (Clavijo et al., 2001; Barros, 2021). Asimismo, es un parámetro que ejerce influencia sobre procesos fisiológicos del crecimiento y desarrollo de las plantas (Lozada y Sentelhes, 2008), considerando que cada especie requiere una temperatura óptima, mínima y máxima para su sobrevivencia y su desarrollo normal, si esta no cuenta con sus límites demandados el crecimiento se ve afectado (Chaves y Gutiérrez, 2017; Gu et al., 2020). La temperatura influye en el crecimiento de las plantas, a temperaturas muy altas y bajas se reprime el creci- miento (Sabehat et al., 1998; Chaves y Gutiérrez, 2017). La precipitación incrementa la disponibilidad de humedad en el ambiente (Martínez et al., 2013), determinando el crecimiento de las formaciones arbóreas (Duval et al., 2015). El bosque La Palma presenta una temperatura entre 12,8 °C y 26,5 °C, con precipitaciones de 613 a 908,7 mm año-1, características que permiten el crecimiento y desarrollo de la especie de C. pubescens.
La pendiente presentó una correlación muy baja (0,02) con la altura de la C. pubescens infiriendo que la pendiente no ejerce influencia directa en la especie. La posición de la pendiente influye en el crecimiento de los árboles al afectar patrones de iluminación solar (Nüchel, 2019; Mašek et al., 2021), distribución de agua y pre- cipitación (Duan et al., 2022).
El pH presentó una correlación muy baja y negativa (-0,14) respecto a la altura del árbol, infiriendo que este parámetro no ejerce influencia significativa en la C. pubescens. La importancia del pH en el suelo radica en el efecto que ejerce sobre las propiedades físicas, químicas y biológicas de este, mediante el intercambio iónico, el pH regula la disponibilidad de macro y micro nutrientes determinando la riqueza de especies (Arocha et al., 2019); la mayoría de nutrientes se encuentran disponibles en pH de 6,5 a 7,5 (Vázquez et al., 2002). Sin embargo, los datos obtenidos de los parámetros edáficos y topográficos evaluados en las parcelas instaladas no presen- taron diferencias considerables, probablemente porque el área de estudio tiene una extensión pequeña.
Análisis de Componentes Principales (ACP) entre las variables edafoclimáticas y topográficas y la altura de árbol del C. pubescens
El ACP permite comparar los individuos entre sí, con el fin de observar patrones y, de esta manera, inferir relaciones entre las variables evaluadas, siempre considerando que, a mayor correspondencia entre las variables, mayor es la capacidad de síntesis del ACP y unos pocos ejes factoriales podrán explicar las variables e individuos eva- luados (Pardo, 2020). La distribución de variables sobre los Componentes Principales 1 y 2 (CP1 y CP2), explicó el 82,0 % de la variabilidad de los datos obtenidos (Fig. 3); el CP1 con una varianza de 51,3 %, indica que SC, pH y Al+3 + H+ refieren una relación en la P2. Mientras que el CP2 refleja el 30,7 % de la variabilidad restante de las variables estudiadas, por su parte, en la P1 se evidencia una estrecha relación entre K, N y DisA. Sin embargo, las variables SaB, Mg+2, Na+, MO, SB, L y Ca+2 muestran una mayor fuerza de asociación (por el menor ángulo que existe entre sus vectores), a nivel de todas las variables del sitio. Las características edáficas presen- taron mayor grado de contribución con respecto a las variables climáticas, indicando que en su conjunto influyen en mayor grado en el desarrollo de la especie. Si se considera que, la influencia de los factores abióticos en la calidad de sitio depende de la escala espacial, por lo general a escala local las variables explicativas son las edáficas (McKenney y Pedlar, 2003). No se han registrado estudios de calidad se sitio de la especie C. pubescens a la fecha; sin embargo, Villar et al. (2018) refieren que, para Cinchona officinalis L. existe una sola calidad de sitio, por lo que no se obtuvo correlaciones fuertes entre la variable dependientes y variables independientes; sin embargo, se observó que el Al y MO presentaron una asociación media de 0,42 y 0,39 respectivamente, infiriendo que, a más presencia de estos, la altura dominante de la especie fue mayor. Reportes de Senilliani et al. (2020) indican que las propiedades edáficas (CE, Ca+2, Mg+2 y Na+) influyeron en mayor cantidad en el crecimiento de plantaciones de Neltuma alba (Griseb.) C.E. Hughes & G.P. Lewis; mientras que el pH influye en menor proporción; y González et al. (2015) señalan que el crecimien- to de las plantas de Jatropha curcas L. var. rufus McVaugh, depende de la fertilidad del suelo. Del mismo modo, Mora et al. (2015) encontraron que el crecimiento de Vochysia guatemalensis Standl, está influenciado por los suelos con pH bajos, bajas concentraciones de Ca+2 y concentraciones medias de Mg+2.
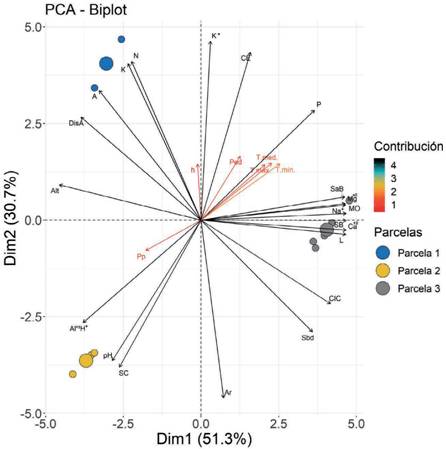
Fig. 3 Biplot de variables edafoclimáticas, topográficos y la altura de árbol del C. pubescens, en el bosque montano La Palma. Fig. 3. Biplot of edaphoclimatic and topographic variables and tree height of C. pubescens in La Palma montane forest.
Caracterización de la calidad de sitio de C. pubescens, en el bosque montano La Palma
El bosque de La Palma donde crece C. pubescens, presenta una temperatura entre 12,8 y 26,5 °C, con precipitaciones de 613 a 908,7 mm año-1, con pendientes de 24 a 48 % y entre los 2 800 msnm a 2 846 msnm. Con respecto al suelo forestal, en el horizonte O, se evidenció una gran presencia de masa radicular, asimismo, en la P2 mostró una profundidad de 12 cm, seguida de P1 (10 cm), P3 (9 cm). Estos datos de profundidad son ligeramente mayores a los reportado por Villar et al. (2018) en suelos del bosque natural de Pagaibamba para C. officinalis L, quienes reportan profundidades de 0 a 10 cm. Con respecto al horizonte “A”, P2 presentó una profundidad de 14,4 cm, seguida de P3 (11 cm) y P1 (6 cm). Estos resultados, evidencian la fragilidad de los suelos en los que crece la especie en el bosque La Palma. Las características obtenidas de la lectura de calicatas nos permitieron realizar la clasificación de parámetros edáficos según el D.S. Nº 005-2022-MIDAGRI (MIDAGRI, 2022). Finalmente, se obtuvo la clasificación de uso mayor para cada parcela evaluada como se detalla en la Tabla 4.
Referente a la textura del suelo, son suelos franco arenosos (Fr. A) coincidiendo con Villar et al. (2018) para suelos de C. officinalis L, quienes refieren que crece en suelos con profundidad media, con textura media a pesada y suelos franco arenosos, por su parte Tara spinosa en bosque naturales también crece y desarrolla en suelos franco arenosos (Villena et al., 2022). Las tres parcelas presentaron un pH fuerte- mente ácido (3,58 - 3,62), característica adecuada para la especie tal como lo reportan Villar et al. (2018). Del mismo modo, Mollinedo et al. (2005), encontraron que la relación suelo-árbol y factores de sitio para Tectona grandis L.f, en los primeros 20 cm de profundidad, en pH = 5,5 el IMA es menor que a un pH >5,5. Sin embargo, se pudo comprobar que no existe una correlación significativa entre h y el pH del suelo, debido a que Cinchona está bien adaptada a suelos ácidos naturales tal como
Tabla 4 Clasificación de parámetros edáficos y clasificación de uso mayor (CUM) del suelo según D.S. Nº 005-2022-MIDAGRI, Perú. Table 4. Classification of soil parameters and classification of major land use (CUM) according to D.S. Nº 005-2022-MIDAGRI, Peru.

riesgo de erosión, c: limitación por clima.ocurre con otras especies que se adaptan a este tipo de suelos, como por ejemplo Pinus taeda L. (Subedi y Fox, 2016). Todas las parcelas presentan un alto contenido de MO, siendo P3 la de mayor contenido 19,31 %, seguida de P1 y P2 con 7,24 % y 5,17 % respectivamente, resultados similares a los reportados por Villar et al. (2018) quienes mencionan que C. officinalis L., requiere suelos con alta a media cantidad de MO, esta característica juega un papel primordial en la fertilidad de los suelos, como fuente de energía para microorganismos y fuente de nutrientes de las plantas, siendo su mayor contribución de la CIC (Sales Dávila, 2006; Cabrales et al., 2011, Ríos et al., 2016). Se registraron niveles medios a altos de P, para la P3 (24 ppm), P1 (21,4 ppm) y P2 (13,1 ppm); superiores a los registrados por Villar et al. (2018) quienes anotan un máximo de 12,40 ppm y un mínimo de 1,90 ppm, permitiendo inferir que algunas especies del género Cinchona se adapta en altos y bajos niveles de P, el cual es esencial para el crecimiento de las plantas (Gueçaimburu et al., 2019), debido a que juega un papel fundamental en el transporte del floema, siendo absorbido por las células, se ve inmediatamente involucrado en procesos metabólicos, translocándose en dirección hacia arriba o hacia debajo de la planta (Mengel y Kirkby, 2000).
El K presente se registra en concentraciones medias, siendo en la P1 (138 ppm),
seguido de P2 y P3 (111 ppm), valores que se encuentran en el rango reportado por Villar et al. (2018); quienes encontraron concentraciones de 105 a 210 ppm. Estos reportes indican que las especies de Cinchona en bosques montanos prosperan en suelos con contenidos medios de aproximadamente 150 ppm de K, como indica Daniel et al. (1982) para especies forestales. Este elemento es vital no solo en rela- ción a su presencia en los tejidos de las plantas, sino que también, a sus funciones fisiológicas y bioquímicas; se ha evidenciado que K se transporta principalmente a los tejidos meristemáticos, y usualmente de los órganos de las plantas adultas es redistribuido a los tejidos más jóvenes (Mengel y Kirkby, 2000; Mattos et al., 2020). Por ejemplo, se registra que la velocidad de traspaso de C en el tronco es dos veces mayor en árboles fertilizados con K (Epron et al., 2015). Asimismo, K presenta un impacto significativo en el crecimiento de los árboles (Fromm, 2010). En individuos de Eucalyptus grandis Hill ex Maiden (W. Hill ex Maiden), K incrementó la síntesis primaria neta aérea en un 100 % mejorando el índice de área foliar (Laclau et al., 2009). En Pinus radiata D. Don, se evidenció una duplicación en la respuesta del crecimiento del tallo (Smethurst et al., 2007).
Los suelos arcillosos presentan una CIC elevada y alta retención de agua que los
suelos no arcillosos (Mengel y Kirkby, 2000). Se reportan valores entre 2 meq/100 g para suelos arenosos a 57,5 meq/100 g en suelos arcillosos (Weil y Brady, 2016). Para este estudio se observó un CIC de mayor concentración en P3 (24,32 meq/100 g), P2 (21,76 meq/100 g) y finalmente la P1 que presenta menor CIC (19,68 meq/100 g), siendo de 10 meq/100 g hasta 20 meq/100 g comunes (Mengel y Kirkby, 2000). Con relación a la suma de cationes cambiables (Ca+2, Mg+2, K+, Na+, Al+3+H+) la P2 presenta mayor cantidad (13,61 meq/100 g), seguido por P3 (11,52 meq/100 g) y P1 (11,38 meq/100 g). El contenido de N en áreas con clima tropical puede variar entre 0,02 y 0,4 % y en suelos ricos en materia orgánica puede llegar hasta 2 % (Robinson, 1994). Se ha reportado dos formas de transporte del N en especies forestales, argini- na (ARG) como principal compuesto de transporte de N en especies de bosques de
sabanas y bosques monzónicos caducifolios y como amido-N en especies de bosques monzónicos perennifolios (Schmidt y Stewart, 1998). Considerándose, en este sentido como el elemento más importante debido a que las especies lo requieren en grandes cantidades; la misma que varía en el suelo en función al clima, localidad y tipo de suelos (Daniel et al., 1982). En el área estudio se encontraron contenidos de N en la P3 y P1 (0,79 y 0,68 %) respectivamente y los niveles más bajos se observa en la P2 (0,60 %), concentraciones altas que aseguran el desarrollo de la especie.
CONFLICTO DE INTERESES
Los autores declaran no tener ningún conflicto de intereses.
CONCLUSIONES
El bosque montano La Palma, presenta una sola clase de sitio (I) óptimo para el crecimiento y desarrollo de C. pubescens. Se evidencia que en el área de estudio las variables edáficas tienen mayor influencia que las variables climáticas sobre el crecimiento de la especie, que requiere de suelos superficiales a moderadamente pro- fundos, con textura Fr. A, fuertemente ácido, con alto contenido de MO, contenido medio a alto de P, medio de K y contenido medio de N, con pendiente entre 38 % a 48 %, micro relieve de ondulado suave a ondulado, erosión muy ligera a ligera; las parcela 1 presentó calidad agronómica media con limitación por suelo, topografía riesgo de erosión y limitación por clima, la parcela 2 y 3 mostró calidad agronómica media con limitaciones en suelo y clima. En ese sentido, las iniciativas de prácticas silvícolas como enriquecimiento de áreas degradadas o macizos de C. pubescens, en el área de influencia del bosque montano La Palma, deberán considerar la realiza- ción de análisis de suelo que permitan verificar los requerimientos mínimos para la especie.

