











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCION
Los procesos tafonómicos pueden incluir una amplia variedad de procesos, siendo uno de los más significativos la alteración por animales carroñeros (Haglund, Reavy y Swindler, 1989). Los animales carroñeros pueden infligir una amplia gama de modificaciones a un cuerpo, y cada especie tiende a modificar los restos de manera específica marcando un cierto patrón conocido comúnmente como "firma tafonómica” (Hannigan, 2015).
La acción de los carroñeros puede destruir completamente huesos enteros o partes de ellos, lo que podría implicar la incapacidad de recuperar las diferentes partes anatómicas y complicar el proceso de identificación personal (Manheim, Listi y Leitner, 2006).
Cada grupo de carroñeros comúnmente llamado gremio (Sorg, 2013) tiene su propio patrón de modificación y dispersión de los restos. Ese patrón, incluye, entre otras características, cuáles son los huesos que más se dispersan, qué partes de esos huesos son generalmente los que tienden a sobrevivir, qué tipos de marcas se manifiestan, entre otros indicadores (Haglund et al., 1989). Uno puede encontrar más de una huella en un conjunto de huesos, dependiendo del entorno y del estadio de descomposición (Sorg, 2013).
Son muchos los autores que han señalado la pérdida de información en aquellos sitios donde han actuado los carroñeros, ya que estos animales tienden comúnmente a consumir, ocultar y dispersar gran parte de los restos esqueléticos (Bright, 2011; Komar y Potter, 2007; Rodríguez y Bass, 1985; Milner y Smith, 1989). Haynes (1982) señala que la sub representación de falanges, costillas, vértebras, carpianos y tarsianos, puede indicar la presencia de cánidos, porque los lobos, perros y coyotes a menudo consumen estos elementos. El mismo autor, afirma que las tasas de recuperación de los huesos de las extremidades superiores oscilan entre el 50% y el 20%, las de las extremidades inferiores entre el 65% y el 42%, y los huesos del esqueleto axial oscilan entre el 73% y el 61%.
Estos porcentajes muestran la preferencia y la facilidad de desarticular elementos esqueléticos específicos por parte de los carroñeros, existiendo una relación positiva entre el porcentaje de un cuerpo recuperado y la identificación, lo que implica que la recuperación exitosa de elementos esqueléticos de una escena forense, afectaría el resultado final de una identificación. Es por este motivo, que un mayor conocimiento sobre el comportamiento de animales carroñeros debería proporcionar una mejor estrategia para la ubicación de los elementos esqueléticos (Haglund, Reavy y Swindler, 1988; Young, Márquez-Grant, Stillman, Smith, y Korstjens, 2015a).
La alteración de un cuerpo por carroñeros en el contexto forense, no es inusual. Durante el 2011, se notificaron en el Reino Unido unos 1.271 casos de personas desaparecidas, donde 267 resultaron en muertes. Del total, un 51,69% se produjo en lugares rurales o periurbanos, donde los cuerpos estaban expuestos a la acción de carroñeros, tanto salvajes como domésticos, que, en mayor o menor grado, modificaron en gran medida los restos (Young et al., 2015a).
La identificación de la acción de los carroñeros, permitirá una estrategia más eficaz para la búsqueda y recuperación de cualquier elemento esquelético que falte y haya sido desarticulado y esparcido por el terreno. Maximizar la recuperación de los restos humanos es crítica al calcular el número de individuos, la identificación del difunto, la evaluación del trauma, el establecimiento de la forma de la muerte, e incluso inferir el intervalo post mortem (IPM) (Manheim et al., 2006; Mann, Bass y Meadow, 1990, Ubelaker y DeGaglia, 2020).
Aunque los estudios de la acción de los carroñeros y sus efectos en los restos humanos son tratados mayormente por los profesionales forenses, muchas de las interpretaciones se basan más en la experiencia del investigador que en registros minuciosos y detallados (García Putnam, 2014).
El objetivo de este artículo es describir el comportamiento de un grupo de tres carroñeros en una zona de humedales en entornos peri urbanos en la Provincia de Buenos Aires. Por medio de la observación de las diferentes especies involucradas, carancho (Caracara plancus), perro doméstico (Canis familiaris) y zorro (Lycalopex gymnocercus), se diagrama una secuencia de modificación de la carcasa, se evalúa el patrón de dispersión de los restos, y se infiere una relación con el intervalo post mortem, extrayendo conclusiones que pueden ser un aporte significativo para la construcción de modelos tafonómicos forenses.
Planteo del problema
El carroñeo es una estrategia de forrajeo en la que tejidos biológicos muertos y en descomposición, conocidos como carroña, se consumen para satisfacer las necesidades nutricionales. Los carroñeros contribuyen eficientemente al ecosistema, acelerando el ciclo de nutrientes, dispersando los nutrientes concentrados y diluyendo los organismos patógenos asociados con la descomposición (Barton, Cunningham, Lindenmayer, y Manning, 2013; Sincerbox y Di Giangi, 2018).
La carroña no sólo es un recurso de alta calidad, sino que requiere relativamente de poco gasto energético en comparación con una estrategia de caza (DeVault, Rhodes, y Shivik, 2003). Debido a estos extraordinarios beneficios energéticos, la mayoría de los vertebrados consumirán carroña si se presenta la oportunidad.
Los carroñeros mamíferos y aviares pueden modificar los restos humanos y así obstaculizar la interpretación del contexto forense. Sin embargo, no todas las especies carroñeras tienen el mismo comportamiento ni están presentes en todos los escenarios. A pesar de la variabilidad, la mayor parte de las investigaciones sobre la acción de los carroñeros en restos humanos se centra en especies de América del Norte (Domínguez-Solera y Domínguez-Rodrigo, 2011; Haglund et al., 1989; Komar y Beattie, 1998; Willey y Snyder, 1989).
Uno de los grupos más estudiados es el orden Carnivora. Estudios sobre este grupo describen el método de consumo y dispersión de las partes anatómicas (Blumenshine, 1988; Camarós et al., 2012; Haglund, 1997; Moraitis y Spiliopoulou, 2010; Young, 2015b). Los carnívoros generalmente consumen la carne y los órganos, pero también modifican los huesos, dejando marcas que podrían incluir desde simples hoyos y surcos hasta fracturas. El daño producido por carnívoros se concentra mayormente en las epífisis de los huesos largos, los procesos espinosos y transversales de las vértebras, los extremos distales de las costillas, las escápulas y la pelvis (Haglund et al.1989). Los restos también pueden ser desarticulados y dispersados distancias que pueden llegar hasta 3,2km, aunque por lo general no superan un promedio de 100 m (Haglund, 1997).
En muchos contextos geográficos peri urbanos los perros domésticos son, junto al zorro y el coyote, los cánidos más grandes con acceso potencial a restos al aire libre, pudiendo producir importantes alteraciones sobre éstos, optando generalmente por la cabeza y el cuello (García Putnam, 2014).
Es cierto que los cánidos son algunos de los carroñeros más estudiados en detrimento de otras especies en una variedad de regiones (véase Jeong, Jantz y Smith, 2014 y Young, Márquez-Grant, Stillman, Smith, y Korstjens, 2015c). Mientras algunos estudios se centran en una categoría taxonómica, otros comprenden múltiples especies, evaluando la interacción entre ellas (Cantú, 2014; Sincerbox y Di Giangi, 2018; Ubelaker y DeGaglia, 2020).
En los últimos años se han realizado estudios tafonómicos forenses sobre la acción de aves carroñeras destacándose los trabajos de Robert y Vigne (2002a, 2002b) sobre Gypaetus barbatus (quebrantahuesos), Laudet y Selva (2005) sobre Corvus corax (cuervo) y Reeves (2009) sobre C. atratus y Cathartes aura (jote cabeza colorada). Algunas observaciones realizadas en el centro de Texas, demostraron que los buitres podían esqueletizar completamente los restos en tan solo 96 horas (Reeves 2009). Aunque estas especies son responsables de la desarticulación parcial y dispersión, lo hacen mucho menos que los cánidos (Kjorlien, Beattie y Peterson, 2009; Marín Arroyo, Fosse y Vigne, 2009).
En Argentina se han realizado estudios similares, destacándose los trabajos sobre alteración de carnívoros sobre restos humanos arqueológicos y forenses (Martin ,2002; Manesegosa, Marchiori y Giannotti, 2020). En cuanto al carroñeo de aves, existen datos para el Caracara plancus (carancho) (Mallet, 2016; Montalvo et al., 2011), Coragyps atratus (jote de cabeza negra) (Ballejo, Fernández y De Santis, 2012), entre otros. La mayor parte de estas especies dejan hoyos y arañazos en individuos adultos y pueden destruir o transportar partes anatómicas en individuos inmaduros o especies de mediano y pequeño porte (Morton y Lord, 2006; Reeves, 2009).
En cuanto a los órdenes Rodentia y Artiodactyla la investigación es más reducida que la investigación sobre Carnivora (Berryman, 2002; Haglund, 1997; Hannigan, 2015).
MATERIAL Y MÉTODOS
Metodología
El término humedales se aplica a una gran variedad de hábitats interiores, costeros y marinos con determinadas características: se trata de extensiones de marismas, pantanos, turberas o superficies cubiertas de agua, de régimen natural o artificial, permanentes o temporales, estancadas o corrientes, dulces o saladas, incluidas las extensiones de agua marina, cuya profundidad en marea baja no exceda los seis metros (RAMSAR, 2006). El agua, que juega un papel fundamental en la definición de la estructura y las funciones ecológicas de los humedales, determina sus características y su variabilidad tanto en el tiempo como en el espacio, afectando definitivamente su diversidad biológica.
En todo el territorio de la Provincia de Buenos Aires, se destaca la presencia de lagunas, permanentes o temporarias de aguas dulces o salobres. La superficie regional de humedales potencial asciende al 47%, sin embargo, de acuerdo con el criterio de cobertura apenas llega al 10% tratándose de un área con una larga historia de actividad agrícola y ganadera cuyo progreso en muchos casos se asoció al desarrollo de obras de infraestructura hidráulicas (Mulvany et al.,2019).
El clima de la zona estudiada, es subtropical-templado, con un gradiente decreciente de humedad y temperatura de Este a Oeste y Sudoeste en sentido latitudinal. La temperatura media registrada para la subregión es de 15,6 °C y la precipitación de 879 mm (Mulvany et al., 2019; Passarelli, Rolleri, Ciciarelli, Dedomenici y González, 2014).
La vegetación acuática o macrófita representa, sin duda, un rasgo sobresaliente de los humedales pampeanos al proporcionar complejidad estructural para el crecimiento, alimentación, reproducción y cría de peces, desarrollo de insectos acuáticos, aves, anfibios, etc. En las lagunas pampeanas es frecuente la presencia de macrófitas emergentes como el junco (Schoenoplectus californicus) y la totora (Typha spp.) (Benzaquen et al., 2017). La Flora autóctona de estos ecosistemas está comprendida por numerosas especies de árboles entre las que se pueden mencionar la tala (Celtis ehrenbergiana), espinillo (Acacia caven), sauce criollo (Salix humboldtiana), ceibo (Erythrina crista-galli), que conforman el típico paisaje bonaerense que va desde la barranca hasta la zona ribereña. A medida que aumenta la presencia de agua se pasa de los pastizales de cortadera, flechilla y chilca a los pajonales con juncales (Schoenoplectus californicus), totorales (Typha spp.), y lirio amarillo (Iris pseudacorus) (Passarelli et al., 2014).
Todo esto conforma el hábitat perfecto para la fauna, presente con gran cantidad de especies de más de un centenar de aves entre las que se observan garzas (Familia Aardeidae), patos (Anatidae), gallaretas, gallinetas, chorlos y gaviotas, y otros grupos de fauna como tortugas, ranas, sapos, lagartos overos, coipos, comadrejas y cuises (ACUMAR, 2006). Esta subregión de humedales incluye áreas protegidas de jurisdicción nacional, provincial y municipal, algunas también designadas Reservas de Biósfera y sitios RAMSAR, que en algunos casos poseen el objetivo de preservar diferentes tipos de humedales (Benzaquen et al., 2017). Una de ellas es La Reserva Natural Bosques de Ciudad Evita , en el Partido de La Matanza, constituyendo una de las áreas protegidas más extensas con cerca de 281 hectáreas (Benzaquen et al., 2017; Kazimierski y Badano, 2019).
El Proyecto Cronos forma parte de un estudio experimental forense llevado adelante por el Área de Criminalística y Estudios Forenses de GNA (IUGNA- ESCUGEN - Ministerio de Seguridad Res. 124.2018). Se desarrolla en un entorno natural de humedales que forman parte de la Cuenca Matanza Riachuelo (2.047km²). Dentro de la mencionada cuenca, se encuentra la Reserva Natural de bosques Ciudad Evita, partido de La Matanza, donde cerca de 15 ha aproximadamente corresponden al predio de la Escuela de Gendarmería Nacional Martín Miguel de Güemes 34° 44´ 25´´ S 58° 31´ 40´´ O (Fig.1).

Figura 1 Cuenca Matanza - Riachuelo y ubicación del área de estudio (Reserva Municipal Ciudad Evita - Escuela de Gendarmería Nacional - Círculo en rojo). Fuente: Instituto Nacional del Agua. ACUMAR
En ese entorno natural se depositaron tres ejemplares de cerdo doméstico (Sus scrofa) (en adelante, Unidades Experimentales UE) con el objeto de monitorear su evolución entomológica, química y tafonómica. Los ejemplares de raza híbrida con un peso de 25kg, fueron provistos por el Bioterio de la Universidad Maimonides siguiendo el protocolo internacional de eutanasia sincrónica (European Comission 1997; LASA/RSPCA 2010) (Fig.2).

La UE 1 fue depositada sobre el suelo decúbito lateral en una zona abierta y descampada. La UE 2 fue depositada en forma similar a 150m al SE, pero dentro de un bosque de acacias y sauces a 3m del borde de la barranca del río Matanza. Ambas UE, se protegieron con jaulas de acero galvanizado de 2x1,5x1,5m con malla de 0,5mm, para evitar que sean alterados por fauna, y se comenzó a relevar secuencialmente la evolución entomológica.
La UE 3 (Fig.3 - 34°44'29.86"S, 58°31'31.92"O), objeto del presente estudio, fue depositada vestida en una zona de matorral intermedia entre el bosque de galería y el descampado, al pie de una acacia y sin estructura de protección, con el objetivo de relevar las alteraciones por carroñeros. Para asegurar la máxima obtención de registros e impedir que los carroñeros puedan arrastrar todo el ejemplar por fuera del área de seguimiento y perder su localización, se sujetó con una cuerda de 2mm x1,20m de un extremo de la pata trasera derecha a una estaca de seguridad. La duración total de la observación para la UE 3 fue de 38 días, tomando como inicio del intervalo post mortem (IPM) la eutanasia (día 0: 10 AM del 14 de septiembre de 2019, hasta la pérdida total de registro óseo el 22 de octubre 14 PM).

La evolución tanatológica de la UE 3, se registró diariamente siguiendo las etapas de descomposición según Galloway (1997) respaldado por un archivo fotográfico obtenido con una cámara Nikon Coolpix B500 y editado mediante Adobe Bridge.
En tanto que el registro tafonómico se llevó a cabo utilizando una ficha de registro diario complementada mediante el uso de una cámara Smart Tech HC 290 M, infrarroja activada por movimiento cada 5 segundos. Mediante este registro video-gráfico automático, se obtuvo: fecha, hora, y duración de cada evento protagonizado por un carroñero a lo largo de todo el estudio. El evento es la unidad de análisis y se refiere a la llegada, interacción y retiro de un carroñero de la UE 3. En este sentido, se registró el orden de acceso de cada animal, el tiempo de interacción, y las modificaciones producidas al cuerpo con el fin de establecer una comprensión clara de cómo estos carroñeros influyen en los cadáveres depositados en la superficie. No existe un rango estándar utilizado para clasificar el tamaño del carroñero, sin embargo, para esta investigación se utilizó el peso para discriminar en: grandes (>40kg), medianos (11-40kg) y pequeños (<10kg) (Delaney-Rivera et al., 2009). Los datos climáticos diarios en intervalos de una hora, se obtuvieron de la estación meteorológica automática del Proyecto Cronos Meteostar.
Para relevar la dispersión y localización de las partes anatómicas, se utilizó un GPS Garmin eTrex Legend. A partir de los datos georreferenciados se confeccionó un mapa de distribución espacial en metros a partir de un centro geográfico que se toma como punto 0 (articulación escapulo humeral), Figura 4. Para la localización de las unidades anatómicas desarticuladas y dispersadas se utilizó, de forma experimental, un Localizador Rastreador Bluetooth Itag 4.0. insertado en la mandíbula inferior. La búsqueda de las restantes partes anatómicas se completó con un método de enlace, similar al método de búsqueda (winthropping), que se basa en ajustar la dirección de búsqueda en función de la identificación de señales o puntos de referencia que conducirán a la recuperación de hallazgos (Adairi y Kolz, 1998; Matuszewski et al., 2019).

Figura 4 Anatomía de Sus scrofa y sus partes anatómicas. 1 (Cráneo y mandíbula), 2 Vértebras cervicales), 3 (Escápula), 4 y 5, 6 (Costillas) 7, 8, 9 y 10 (Patas delanteras), 11, 12, 13, 14, 15 y 16 (Patas traseras), 17, 18, 19 y 20 (Pelvis, lumbares, caudales). Punto 0 a partir del cual se mide la desarticulación
Se confeccionaron las áreas de dispersión utilizando Map Maker, mediante la confección de polígonos de dispersión para cada jornada de mapeo. Las partes anatómicas para UE 3 se tomaron individualmente como 206 elementos esqueléticos. Todos los cómputos se presentan como totales de cada categoría anatómica independientemente de si se recupera como fragmentado o como partes anatómicas articuladas.
Por último, se utilizó el paquete estadístico para datos circulares Oriana 4.0. para evaluar la distribución direccional (desviación estándar) representada como un círculo alrededor del centro mostrando el grado de concentración o dispersión esquelética mapeada. Mediante Info Stat se realizó una regresión logística binomial para evaluar la relación entre el IPM y la dispersión de los elementos esqueléticos.
Estudio del material
Tres especies de animales carroñeros interactuaron con la UE 3, carancho (Caracara plancus), perro (Canis familiaris) y zorro pampeano (Lycalopex gymnocercus). El género Caracara, tiene una distribución limitada en América del Norte, pero está presente desde México hasta Tierra del Fuego (Mallet, 2016). El Caracará Del Sur, conocido vulgarmente como Carancho (Caracara plancus), es un raptor común en Argentina, y su hábitat son los campos abiertos, pastizales naturales, matorrales, pantanos, pastos, humedales y áreas agrícolas (Canevari et al., 1991).
El Caracará del Sur es típicamente un ave rapaz oportunista, aunque pocos estudios cuantitativos de su dieta están disponibles (Balza, 2016), gran parte de su alimentación es la carroña (animales muertos en carretera, cadáveres de ganado). Sin embargo, se lo encuentra cerca de mataderos y vertederos de basura (Mallet, 2016), mientras que en la Patagonia esta especie se asocia con carroña de ganado (Travaini, Donazar, Ceballos e Hiraldo, 2001).
En cuanto al perro, pese a la gran diversidad de razas, formas y tamaños, todos son miembros de la misma especie: Canis familiaris. Aunque su forma de vida sea doméstica, están emparentados con los lobos, los zorros y los chacales (Sillero-Zubiri, Hoffman y McDonald, 2004). Los Canidae contemporáneos son la familia más extendida del orden Carnivora existente, con al menos una especie presente en todos los continentes excepto en la Antártida. En el último siglo los rangos geográficos de siete especies han aumentado, ocho han disminuido y nueve se han mantenido estables (Macdonald y Sillero-Zubiri, 2004).
Los perros cimarrones o asilvestrados tienen origen en los perros domésticos. Estos canes que, por variadas circunstancias se alejan de la presencia humana, producen a menudo daños económicos a la propiedad y a las personas, a veces de consideración, frecuentando los alrededores del casco urbano (basurales) o inclusive extendiéndose a las explotaciones rurales. En las 15 hectáreas del predio perteneciente a la Escuela de Gendarmería Nacional Martín Miguel de Güemes, conviven jaurías (entre seis y diez) de canes asilvestrados, que frecuentemente extienden su territorio hasta la Reserva Municipal lindante. Se han registrado al menos dos cubiles en las cercanías de las instalaciones de la Institución y también merodeando sobre los desperdicios del servicio de cocina.
En cuanto al zorro pampeano Lycalopex gymnocercus (Birochio, 2008), es una de las especies pertenecientes al género Pseudalopex, que, pese a su amplia distribución, ha sido muy poco estudiada. La información sobre su distribución y sobre las áreas de solapamiento con el zorro gris P. griseus resulta insuficiente (Lucherini, Pessino y Farias, 2004), la mayor parte de la información se encuentra en un trabajo de Crespo (1971).
Es un cánido de mediano tamaño, a menudo confundido con P. griseus en el límite sur de su distribución y con el zorro de monte Cerdocyon thous en la zona norte de su distribución (Lucherini et al., 2004). En la provincia de Buenos Aires se determinó que su alimentación incluye roedores y aves, pero también insectos, frutos y fundamentalmente carroña (Castillo, 2003 Farías, 2000). Particularmente, Birochio (2008) halló como parte de sus hábitos generalistas y oportunistas que la carroña de caballo cimarrón constituye un importante ítem alimenticio, teniendo bajo impacto como predador de ganado doméstico.
Aunque son carroñeros solitarios, se caracterizan por su gran movilidad, llegando a desplazarse hasta 2,5ha diarias. Es una especie que puede adaptarse fácilmente a distintos ambientes lo que le ha permitido permanecer dentro de los agro ecosistemas (Birochio, 2008; Lucherini et al., 2004).
El primer registro de la presencia de un carroñero sobre la UE 3, fue un ejemplar de Caracara plancus (carancho) el día 1 de IPM a las 11.21, quien continuó visitando regularmente el sitio durante 22 días (Fig. 5). El carancho actuó primero sobre el hocico (lengua) y los ojos. También sobre el cuello y las mandíbulas. Alternativamente avanza y picotea sobre la pata trasera. Se pudo apreciar una hendidura de 7cm x 2cm de profundidad con bordes lisos sobre el muslo externo de la pata trasera izquierda, compatible con elemento filoso (garra). En los eventos sucesivos dentro del primer día de contacto, el carancho logra acceder al vientre y como consecuencia comienza a devorar parte de las entrañas.

Al concluir cada evento, luego de alimentarse, transporta carroña a un nido localizado sobre un árbol de eucaliptus a 130m al NE. Sólo en una oportunidad se presentó simultáneamente un segundo ejemplar a los 8 días de IPM, no participando del carroñeo. Una vez producido el acceso al vientre, comienza a avanzar sobre la región del tórax, aunque la ropa dificulta el acceso a esta zona anatómica (Young, 2013).
El carancho actúa en 37 eventos a lo largo de 38 días de IPM, con una duración total de 580 minutos y un promedio de 16 minutos por evento (Tabla 1). Otras aves muy comunes como el benteveo (Pitangus sulphuratus) y gorrión común (Passer domesticus) visitaban en grupos de dos a tres individuos la UE 3 regularmente en los lapsos libres de competidores, mayormente entre las 10 y 16hs, dando cuenta de las larvas que se criaban sobre la carroña.
TABLA 1 Registro de eventos para Caracara plancus (IPM: Intervalo Post Mortem)
| Día | IPM | Número de | Promedio | Total Eventos |
| Mes | Eventos | en minutos | en minutos | |
| 14-sep | 0 | 0,0 | 0,0 | 0,0 |
| 15-sep | 1 | 5 | 38 | 190 |
| 16-sep | 2 | 4 | 15 | 60 |
| 17-sep | 3 | 5 | 29 | 145 |
| 18-sep | 4 | 5 | 10 | 50 |
| 19-sep | 5 | 8 | 9,75 | 78 |
| 23-sep | 9 | 2 | 11 | 22 |
| 24-sep | 10 | 2 | 7.5 | 15 |
| 30-sep | 16 | 0,0 | 0,0 | 0,0 |
| 1-oct | 17 | 1 | 6 | 6 |
| 2-oct | 18 | 0,0 | 0,0 | 0,0 |
| 3-oct | 19 | 0,0 | 0,0 | 0,0 |
| 4-oct | 20 | 1 | 1 | 1 |
| 5-oct | 21 | 3 | 3,33 | 10 |
| 6-oct | 22 | 1 | 3 | 3 |
| 7-oct | 23 | 0,0 | 0,0 | 0,0 |
| 10-oct | 26 | 0,0 | 0,0 | 0,0 |
| 22-oct | 38 | 0,0 | 0,0 | 0,0 |
| Día | 37 | 163,55 | 580 |
El día 5 de IPM, a las 9.09 AM la cámara de detección de movimiento capturó imágenes de tres ejemplares de Canis familiaris merodeando la UE 3 durante los primeros minutos y accediendo después al tórax a partir de la abertura del vientre realizada por el carancho (Fig. 6). Fragmentos proximales de costillas presentan huellas de acceso preferencial a esta zona. Los perros son los únicos que actúan en grupo (entre dos a cuatro ejemplares) pero accediendo a la carroña como máximo de a dos (2) individuos, sin evidencia de competencia intra-específica.

El día 9 de IPM, la UE 3 se encuentra partida en dos a nivel de primeras dorsales y sólo se encuentran los dos cuartos unidos mediante tejido conectivo (Cuero). Entre el día 17 y 18, los perros acceden al cuarto delantero junto con la cabeza y lo transportan fuera del sitio el día 19 de IPM. La dispersión de la cabeza por un lado y cuarto delantero por el otro, continúa el día 21 y concluye el día 38 de IPM. En resumen, esta especie actúa en forma irregular (pulsos) sobre la UE 3 durante 13 eventos con una duración total de 56 minutos a lo largo de 38 días de IPM, a un promedio de 4,3 minutos de duración por evento (Tabla 2).
TABLA 2 Registro de eventos para Canis familiaris
| Día | IPM | Número de | Promedio | Total Eventos |
| Mes | Eventos | en minutos | en minutos | |
| 14-sep | 0 | 0,0 | 0,0 | 0,0 |
| 15-sep | 1 | 0,0 | 0,0 | 0,0 |
| 16-sep | 2 | 0,0 | 0,0 | 0,0 |
| 17-sep | 3 | 0,0 | 0,0 | 0,0 |
| 18-sep | 4 | 0,0 | 0,0 | 0,0 |
| 19-sep | 5 | 1 | 13 | 13 |
| 23-sep | 9 | 5 | 8,75 | 35 |
| 24-sep | 10 | 0,0 | 0,0 | 0,0 |
| 30-sep | 16 | 0,0 | 0,0 | 0,0 |
| 1-oct | 17 | 1 | 8 | 8 |
| 2-oct | 18 | 1 | 0,0 | 0,0 |
| 3-oct | 19 | 1 | 0,0 | 0,0 |
| 4-oct | 20 | 0,0 | 0,0 | 0,0 |
| 5-oct | 21 | 1 | 0,0 | 0,0 |
| 6-oct | 22 | 1 | 0,0 | 0,0 |
| 7-oct | 23 | 0,0 | 0,0 | 0,0 |
| 10-oct | 26 | 1 | 0,0 | 0,0 |
| 22-oct | 38 | 1 | 0,0 | 0,0 |
| 13 | 29,75 | 56 |
La UE 3 fue visitada el día 16 de IPM por un ejemplar de zorro (Lycalopex gymnocercus), (Fig. 7), logrando llevarse la pata trasera izquierda (la que no estaba unida a la estaca de seguridad). El día 22 y 23 de IPM, accede nuevamente y se lleva la pata trasera derecha. La presencia del zorro en el sitio cuenta con una duración total de 12 minutos, a lo largo de 3 eventos y un promedio de 4 minutos de duración por evento (Tabla 3). Se observó, al igual que en otros estudios, que el zorro interactúa con la UE 3 en varias etapas, acercándose más en cada una de ellas, concentrándose mayormente en las extremidades y no en la cabeza ni el tórax (Young et al., 2015a).

TABLA 3 Registro para Lycalopex gymnocercus
| Día | IPM | Número de | Promedio | Total Eventos |
| Mes | eventos | en minutos | en minutos | |
| 14-sep | 0 | 0,0 | 0,0 | 0,0 |
| 15-sep | 1 | 0,0 | 0,0 | 0,0 |
| 16-sep | 2 | 0,0 | 0,0 | 0,0 |
| 17-sep | 3 | 0,0 | 0,0 | 0,0 |
| 18-sep | 4 | 0,0 | 0,0 | 0,0 |
| 19-sep | 5 | 0,0 | 0,0 | 0,0 |
| 23-sep | 9 | 0,0 | 0,0 | 0,0 |
| 24-sep | 10 | 0,0 | 0,0 | 0,0 |
| 30-sep | 16 | 1 | 4 | 4 |
| 1-oct | 17 | 0,0 | 0,0 | 0,0 |
| 2-oct | 18 | 0,0 | 0,0 | 0,0 |
| 3-oct | 19 | 0,0 | 0,0 | 0,0 |
| 4-oct | 20 | 0,0 | 0,0 | 0,0 |
| 5-oct | 21 | 0,0 | 0,0 | 0,0 |
| 6-oct | 22 | 1 | 5 | 5 |
| 7-oct | 23 | 1 | 3 | 3 |
| 10-oct | 26 | 0,0 | 0,0 | 0,0 |
| 22 0ct | 38 | 0,0 | 0,0 | 0,0 |
| 3 | 12 | 12 |
La Tabla 4, resume el número de eventos y el tiempo de interacción entre las tres especies. La acción del carancho es la más significativa tomando en cuenta el índice de interacción, presentando una correlación negativa con el índice de desarticulación (r: -0,75). Por el contrario, el índice de interacción de los perros es tres veces menor al del carancho, sin embargo, la desarticulación es relevante. El zorro obtiene el mismo índice de interacción de los perros y la mitad del índice de desarticulación. Estos dos últimos carroñeros de tamaño mediano (11 a 40kg), interactuaron mucho menos, tanto en número de eventos como el tiempo con UE 3, provocando, sin embargo, una alteración significativa sobre los restos. Estimamos que las diferencias temporales en la detección de la UE 3 por los carroñeros no tienen implicancias con la duración de los eventos y la alteración de los restos. En efecto, el carancho llegó 24 horas después de depositada la UE 3 (1 día de IPM), los canes llegan a las 120 horas (5 días de IPM), y el zorro a las 384 horas (16 días de IPM).
TABLA 4 IE: Índice de Interacción (minutos totales/número de eventos). ID: Índice de desarticulación (huesos desarticulados/total de huesos)
| Especie | Eventos | Minutos | ID | IE | Peso en Kg |
| Caracara | 39 | 557 | 0,0048 | 14,28 | 1,6 |
| Canis | 9 | 75 | 0,325 | 8,33 | 37 |
| Lycalopex | 3 | 12 | 0,15 | 4 | 14 |
La Figura 8 muestra la sucesión de eventos por día de observación y su duración en minutos para los tres carroñeros. Es notoria la presencia del carancho (eventos) a lo largo de todo el periodo, sin embargo, no constituye una especie que destruya o transporte restos esqueléticos fuera del área de depositación original. Le siguen en frecuencia de eventos los perros en dos periodos y por último una breve aparición del zorro hacia el final del mismo.

RESULTADOS Y DISCUSIÓN
Desde hace décadas los ecólogos han estudiado la organización de las comunidades animales y los factores que influyen sobre su comportamiento (Sincerbox y Di Gangi, 2018). Sin embargo, muy poca investigación en el área forense involucra información etológica sobre las especies que interactúan y este es un factor crítico para identificar y comprender las alteraciones tafonómicas de los restos implicados (Hannigan, 2015).
La carroña se presenta como un recurso limitado, es decir está disponible sólo inmediatamente después de que un organismo muere, agotándose rápidamente. Además, la carroña está dispersa en parches aislados a lo largo de todo un territorio, lo que asegura que este recurso sólo sea comestible por un tiempo crítico después de la muerte (Sincerbox y Di Giangi, 2018).
La mayoría de los carnívoros se distribuye sobre el territorio como un individuo o en grupos, definiendo el espacio como una porción del área de acción (“home range”) dentro de la cual, un individuo, o un grupo accede a los recursos (Burt, 1943; Maher y Lott, 1995). En consecuencia, el mantenimiento de un territorio puede ser visto como una adaptación a la defensa de un recurso crítico que con frecuencia es el alimento (Maher y Lott, 2000). Dado que la territorialidad involucra tanto beneficios como costos, se espera que los territorios sean del tamaño mínimo necesario para satisfacer los requerimientos de los ocupantes. Un aumento en el tamaño de los territorios podría aparejar costos sin un beneficio neto, sobre todo a causa del incremento de la competencia entre las diferentes especies (Houston, Sinclair y Norton-Griffiths, 1979; Carr y Macdonald, 1986).
Aunque en otros contextos, aves rapaces como los buitres pueden esqueletizar completamente un cerdo adulto en un período de tres horas, a cinco días, el carancho, debido principalmente a su tamaño, y a su comportamiento (no grupal), consume el cadáver básicamente a través del picoteo y el desgarro (Oliver y Graham, 1994; Pokines y Baker, 2013). Como mencionamos antes el carancho abrió rápidamente el vientre exponiendo las vísceras y el tracto digestivo. El tiempo transcurrido desde el primer contacto con la carroña, hasta la apertura del vientre fue de 20 minutos durante el tercer evento del día 1 de IPM.
El carancho transportaba mayormente jirones musculares, vísceras y fragmentos de piel, semejante a lo reportado por otros autores (Traviani et al., 1998). Luego de cada evento, este carroñero volaba en la misma dirección, lo que sugiere que posiblemente estaba suministrando recursos al nido, ya que la temporada de cría es entre junio y diciembre (Traviani et al., 1998; Vargas y Marco Favero, 2007).
En cuanto al zorro, estudios realizados sobre su población, demuestra que la secuencia de acceso a la carroña comienza por las extremidades traseras a las delanteras, seguida por el tórax que incluye la desarticulación y la dispersión de vértebras, costillas, esternón y escápulas (Carson, Stefan y Powell, 2000; García Putnam, 2014). En el caso de la UE 3, el zorro sólo pudo acceder y transportar al cuarto trasero libre de la atadura en el primer evento (día 16 IPM) y la pata trasera restante el 23 de IPM.
Se sabe que, en otros contextos ecológicos, los zorros comienzan a consumir un ciervo después de un tiempo de exposición de 20.4 días (Adair y Kolz, 1998; Jones, 2011; Young et al., 2015b). La UE 3 fue visitada por el zorro el día 16 de IPM e interactúa hasta el día 38 de IMP.
La presencia de Canis, con un peso promedio de 37kg para un ejemplar adulto, indudablemente podría afectar el comportamiento del carancho y del zorro, sobre todo limitar a este último al horario nocturno. En ambientes donde el zorro no es el carroñero más grande (14kg promedio para macho adulto), su acceso a la carroña está restringido por la presencia de cánidos más grandes, como perros, coyotes y lobos, que tienen acceso privilegiado frente a los de menor tamaño, mostrándose agresivos sobre la territorialidad de los restos (García Putnam, 2014; Young et al., 2015a).
La Tabla 5 detalla los horarios de acceso de las tres especies en relación a la escala lumínica estacional. El Caracara posee el rango horario de interacción más amplio dentro del cual ocurren los eventos, entre el amanecer y el atardecer (100%).
TABLA 5 Actividad lumínica para las tres especies de carroñeros.
| Día | IPM | Salida Sol | Puesta Sol | Duración hs día | Caracara | Canis | Lycalopex |
| 14-Sep | 0 | 06.55 | 18.43 | 11.47 | |||
| 15-Sep | 1 | 06.55 | 18.43 | 11.47 | 11.30 a 18.45 | ||
| 16-Sep | 2 | 06.52 | 18.44 | 11.51 | 06.59 a 10.00 | ||
| 17-Sep | 3 | 06.51 | 18.45 | 11.53 | 06.39 a 18.02 | ||
| 18-Sep | 4 | 06.49 | 18.45 | 11.55 | 06.31 a 10.17 | ||
| 19-Sep | 5 | 06.48 | 18.46 | 11.58 | 06.40 a 14.20 | 09.09 a 09.22 | |
| 23-Sep | 9 | 06.42 | 18.49 | 12.06 | 13.00 a 17.50 | 18.23 a 20.21 | |
| 24-Sep | 10 | 06.41 | 18.50 | 12.08 | 07.47 a 14.02 | ||
| 30-Sep | 16 | 06.30 | 18.56 | 12.15 | 22.32 | ||
| 1-Oct | 17 | 06.31 | 18.55 | 12.23 | 16.24 a 18.06 | 17.37 | |
| 2-Oct | 18 | 06.27 | 18.57 | 12.30 | a 11.48 | 19 | |
| 3-Oct | 19 | 06.26 | 18.58 | 12.32 | 09.47 a 15.47 | 02.05 | |
| 4-Oct | 20 | 06.24 | 18.59 | 12.34 | 11.29 a 11.32 | ||
| 5-Oct | 21 | 06.21 | 19.01 | 12.35 | 09 a 15.47 | 18.30 | |
| 6-Oct | 22 | 06.20 | 19.02 | 12.37 | 11.20 a 11.30 | 19 | 02.02 |
| 7-Oct | 23 | 06.18 | 19.03 | 12.38 | 01.16 | ||
| 10-Oct | 26 | 06.16 | 19.05 | 12.40 | 9.30 | ||
| 22 0ct | 38 | 06.14 | 19.06 | 12.42 | 18.45 |
Fuente: Servicio de Hidrología Naval. http://www.hidro.gov.ar/observatorio/resol.asp)
Para Canis familiaris, el registro demostró que los eventos de esta especie se concentran mayormente después del atardecer (78%) entre las 18 y las 20hs. De un total de 13 eventos, en una sola oportunidad se produjo a la mañana, una a la madrugada y el resto al atardecer.
En el caso de Lycalopex en 3 eventos con una duración total de 12 minutos, se dan entre la 1 y 2 AM. Como vimos, sólo se relevó la presencia de un solo ejemplar accediendo a UE 3. En el ciclo de actividad diurna, se observa una disminución en la actividad (por debajo del 30%) entre las 9hs y las 17hs, siendo el período de mayor actividad (más del 70%) entre las 20hs y las 5hs. Al dividir el día en períodos, la actividad máxima se registró durante la noche (68,4%) y la mínima en el período de día (28%).
La acción de Canis expone áreas de tejido blando que podrían ser inaccesibles, tanto para el carancho como para el zorro. En efecto, la ropa constituye una barrera que no restringe, pero que evidentemente dificulta la acción de los carnívoros de menor tamaño. Por lo que esperamos que el daño causado por zorros en tales entornos se podría limitar principalmente a los elementos descartados por los perros.
Es lógico argumentar que donde el zorro es el carroñero más común, la acción sobre la carroña suele tener un patrón diferente que en entornos donde la competencia es más intensa (Young et al. 2015c). A diferencia del perro, el zorro generalmente no suele estar asociado a una desarticulación total, sino a una desarticulación parcial seguida de una secuencia de visitas y de re dispersión de elementos esqueléticos (Birochio, 2008; Young, 2013).
En la Figura 9 vemos el índice de desarticulación a lo largo del período bajo estudio. Las flechas indican la incidencia de perro a los 9 y 17 días de IPM. El zorro interactúa por primera vez el día 16 de IPM, transportando los últimos restos de la UE 3 el día 32 IPM. Como vemos, Canis y Lycalopex son las especies que más alteran los restos de la UE 3.

Figura 9 Índice de desarticulación en todo el período ( ID: Índice de desarticulación (huesos desarticulados/total de huesos)
Aunque no hay agresión entre los gremios, existiría cierta jerarquía que se expresa en la secuencia de acceso de cada especie: Canis desplaza al Caracara y a Lycalopex, el Caracara se presenta ante la ausencia de Canis y Lycalopex, y este último accede cuando no está presente Canis. Por último, los pájaros comen larvas cuando no están presentes los carroñeros (Travaini, Donazar, Rodríguez y Ceballos, 1998).
Es cierto que el comportamiento hacia la carroña por parte de las tres especies, podría estar atenuado por la presencia de acumulaciones de desperdicios orgánicos generados por acción antrópica (Cocina de la Institución y basural general). En este sentido, se han identificado dos cubiles de Canis con crías en las cercanías de estos recursos. Este argumento es interesante, sobre todo en la evaluación de la competencia inter-específica en relación a la UE 3, ya que, en este contexto ecológico de área peri urbana, las alteraciones inferidas por los carroñeros sobre la UE 3, podrían ser consideradas como de valores mínimos (Schultz y Mitchell, 2018).
En efecto, este ecosistema contaría con recursos más concentrados, lo que podría resultar en una alimentación menos agresiva por parte de los carroñeros (Reeves, 2009). Podemos argumentar, en consecuencia, que tanto la actividad de los carroñeros, como la competencia inter gremial es poco significativa, debido en parte, al acceso a las acumulaciones de desperdicios y a que las poblaciones de especies silvestres son reducidas (Vidal Luengos, 2009).
En este sentido, observamos que la actividad de los carroñeros también se ve disminuida por fenómenos climáticos y estos son factores que estarían implicados en el comportamiento de los carroñeros. La Figura 10 muestra los eventos para las tres especies durante el periodo de observación. Es interesante notar que, durante los días en que las precipitaciones son más abundantes, la acción sobre la UE 3 por parte de los carroñeros, disminuye sensiblemente (prueba t bilateral 0.037 valor p < 0,05).
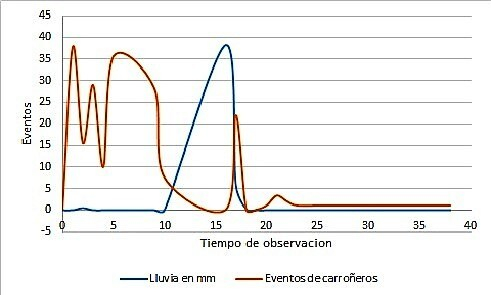
Figura 10 Diagrama de actividad (eventos) de las tres especies sobre la UE3 a lo largo del periodo de observación en relación al nivel de lluvias
En el caso de la UE 3 (Tabla 6), la cabeza, vértebras cervicales y primeras torácicas junto las extremidades anteriores aún articuladas fueron desplazadas por Canis 26m en dirección (NE) entre el día 17 y 19 de IPM. Una pata delantera es desplazada por Canis más de 30m hacia la dirección NE a los 26 días de IPM. Finalmente se observan en el día 38 de IPM, algunas vértebras y fragmentos de costillas a 25m en la misma dirección. Las prendas, completas, fueron finalmente abandonadas a unos 2m, la camisa en dirección (NE) y los pantalones al (SO) el día 26 de IPM. El miembro trasero izquierdo fue desplazado 8m por Lycalopex en dirección (SO) el día 16 de IPM. Finalmente transportó la pata trasera derecha en la misma dirección el día 22 de IPM.
TABLA 6 Dispersión de partes anatómicas por día de IPM de la UE 3.
| Fecha | Parte anatómica | Perímetro | Dispersión en m2 | Distancia lineal en m |
| 1-Oct | A | 1.83 | 0.01 | 2.84 |
| 2-Oct | A | 26.21 | 17.48 | 12.41 |
| 3-Oct | A | 55.86 | 101.46 | 25.86 |
| 7-Oct | B | 17.69 | 15.01 | 8.4 |
| 10-Oct | C | 64.79 | 135.43 | 30.62 |
| 22-Oct | D | 69.78 | 234.83 | 25.43 |
| Total | 236.16 | 504.22 | 105.56 |
La distancia lineal máxima de dispersión de una pieza anatómica (húmero) recuperada fue de 31m, con una distancia promedio para todas las piezas anatómicas de 16,11m. (SD = 11,21), mientras que el tamaño del área total de dispersión de las partes anatómicas superó los 500m². La Figura 11 muestra la dispersión de las partes anatómicas identificadas por elipses superpuestas generadas cronológicamente. Se observa un incremento de la dispersión desde el día 17 hasta el día 38 de IPM, a partir del cual los límites se vuelven difusos debido a la pérdida de elementos esqueléticos. Sostenemos que las distancias de dispersión podrían correlacionarse positivamente con el IPM (Haglund, 1997; Kjorlien et al., 2009), ya que existiría una relación logarítmica entre el incremento del IPM y la dispersión en m². Aunque esta relación debe utilizarse con precaución, el resultado de la regresión lineal (r 0,84), apoyaría la hipótesis de la existencia de tendencias generales en la dispersión de los restos a lo largo del tiempo (Fig. 12).


Figura 12 Línea de tendencia logarítmica y regresión lineal para el IPM y la dispersión en m² a lo largo del tiempo
Si bien algunos estudios han identificado tendencias direccionales significativas para la dispersión de elementos anatómicos (Jones, 2011; Manhein et al., 2006), podemos argumentar que la UE 3 muestra cierta tendencia direccional de dispersión. Los modelos de elipse indicarían una dispersión de los restos en forma no aleatoria a partir del Punto 0. La inferencia para una sola muestra como el estadístico R de Rayleigh, y el test de bondad de ajuste de K de Kuiper (p < 0,005), apoyaría el argumento de que la dirección de la dispersión no es regular, sino de tendencia bi direccional coincidiendo en este sentido, con movimientos hacia áreas de vegetación alta y espesa en el caso del zorro (SO) y en mayor número hacia el cubil en el caso de los canes (NE) indicada con la Moda angular (Fig.13).

Figura 13 Diagrama direccional de dispersión de elementos anatómicos por carroñeros de Sus scrofa en un periodo máximo de 38 días de IPM mostrando la Moda Angular o tendencia direccional (en rojo)
Debemos, en este caso, tomar en consideración que los cánidos como el coyote (Canis latrans), lobo (Canis lupus) y perro doméstico (Canis familiaris) producen, en general, efectos similares. Comienzan mayormente con los tejidos blandos en la zona con presencia de trauma, continuando con la cabeza y el cuello. Pero también producen desarticulación y dispersión sobre los miembros superiores, inferiores y columna vertebral (Haglund et al., 1989).
Sin embargo, el patrón de dispersión del zorro difiere de la de los cánidos más grandes, como el perro doméstico, el lobo y el coyote (Young et al., 2015a, 2017). El zorro accede en múltiples visitas y realiza mordeduras a los restos, intentando transportarlos del sitio de depósito hacia la madriguera. Generalmente accede a las extremidades seguidas por la cavidad torácica incluyendo vértebras y el cráneo. Al ser un carroñero solitario, el zorro intentará dar cuenta del recurso disponible transportándolo a un área más apartada con el objeto de consumir y almacenar recursos sin la amenaza de otros carroñeros (Young et al., 2015b).
Las distancias de dispersión de restos humanos por zorros son similares, pero pueden ser afectados por la condición, la disposición del cuerpo, y la presencia de ropa (Young et al., 2015a). Las distancias de dispersión de carroña y restos humanos realizados por zorros, están entre los 10 y 45 m, transportándolo a áreas de vegetación alta con cubierta densa de árboles o árboles caídos, aunque se ha registrado variaciones que van desde 18 ha en áreas urbanas del Reino Unido (Baker et al., 2000), hasta 1611ha en la tundra canadiense (Jones, 2011; Young et al., 2015a).
Existe un único trabajo sobre la organización espacial del zorro pampeano (Lucherini et al., 2004); en el mismo se obtuvieron tamaños de área de acción entre 73 y 207ha. Aunque los resultados obtenidos para la UE 3 son mucho menores, brindaría una evidencia concreta de la gran flexibilidad ecológica de esta especie teniendo en cuenta que la dieta omnívora y la época de reproducción (primavera y verano) produciría una reducción del área de acción (Birochio, 2008; Vidal Luengos, 2009).
En cuanto a la dispersión de restos de carroña por parte del carancho, vemos que es una especie en donde sus nidos se disponen generalmente en el límite de los parches de recursos (Balza, 2016). En sistemas donde el recurso alimenticio se dispone de forma abundante y predecible, es esperable que las parejas reproductivas no superpongan sus territorios (Balza, 2016). En el caso de la UE 3 la anidada está ubicada en un área de transición (ecotono) entre una reserva natural y borde peri urbano con acceso a los vertederos de basura.
Es particularmente interesante notar que, el carancho no pudo desplazar las prendas, con excepción de liberar el abdomen. Pese a lo limitado del transporte de piezas anatómicas al nido, la acción de esta especie, no debe de ser subestimada. Algunas investigaciones han reportado aves carroñeras como los buitres que despojan de tejidos blandos a un cadáver humano en aproximadamente 5hs (Gretchen, Dabbs y Martin, 2013; Spradley, Hamilton y Giordano, 2012). Es cierto que esta última especie suele presentarse en grupo a diferencia del carancho, sin embargo, todas ellas tienen cierta ventaja frente a los cánidos y otros carroñeros terrestres debido a la combinación de su poderosa vista y su capacidad de volar, por lo que no sorprende su temprana detección de la carroña (Gretchen et al., 2013).
La zona de acción de un animal incluye el área en la que viaja regularmente para realizar distintas actividades como la caza y el apareamiento (Jewell, 1966). Por lo tanto, los tamaños de los rangos de movilidad pueden ser utilizados como un factor crítico al delinear un área de búsqueda en la escena del crimen. La Figura 14 muestra el rango teórico mínimo de radio de acción para cada una de las especies implicadas en este estudio. El rango mínimo se infiere del radio que une la UE 3 con el cubil de los cánidos, localizado en las cercanías del vertedero de la institución, el nido en el caso del carancho y el centro geográfico teórico de la reserva ecológica en el caso del zorro. La suma de los territorios cubre una extensión de 35Ha y esto nos da una idea del rango de acción potencial de estos carroñeros.

CONCLUSIONES
Si bien sabemos que los perros son los animales carroñeros que se reportan con mayor frecuencia en el contexto forense, sobre todo en zonas urbanas y peri urbanas, algunos animales silvestres como el zorro y aves rapaces como el carancho, pueden tener un impacto significativo en la alteración de restos biológicos.
Contamos con la evidencia que, a lo largo de un periodo de 38 días, tres especies de carroñeros acceden en forma secuencial a los restos biológicos de un cerdo vestido de 25kg en los humedales de la Provincia de Buenos Aires, entre los meses de septiembre y octubre. La secuencia de acceso a los restos comienza con el carancho el día 1 de IPM, seguido por el perro (día 5 de IPM) y, por último, el zorro (día 16 de IPM).
En cuanto al perro, su composición gregaria, su mayor tamaño y su superioridad en términos de competencia inter especie local, lo convierte en el candidato que más altera, desarticula y dispersa los restos. Sin embargo, el comportamiento de esta especie en relación a la presencia humana, debe evaluarse, sobre todo con respecto al acceso y disponibilidad de recursos como los vertederos. En cuanto al carancho y al zorro, su capacidad adaptativa en entornos peri urbanos hace que la carroña constituya uno de sus principales recursos, sobre todo en entornos ecológicos con intrusión antrópica.
Advertimos también, que no existiría una relación significativa entre la cantidad de eventos por especie en interacción con la carroña y los procesos de desarticulación y dispersión. El perro y el zorro, son las especies que menos tiempo interactúan con la UE 3, sin embargo, son los principales agentes de alteración anatómica.
Algunos fenómenos climáticos, incidieron en la frecuencia de eventos para las tres especies. En este último sentido, la precipitación fue un factor climático que disminuyó la presencia de los carroñeros y que afectaría indirectamente, el número de eventos, la alteración y la dispersión, ya que los animales no estuvieron activos los días en que la lluvia era intensiva (40mm máximo registrado en el periodo de observación) o en el día inmediato posterior. Por otro lado, la cantidad de cobertura vegetal fue un factor determinante en la dirección en la que se dispersaron las diferentes partes anatómicas, ya que el sitio con menos cobertura hacia el (NE) fue utilizado por los perros, mientras que el área que estaba cubierta por árboles y matorrales densos, de difícil acceso (SO) fue utilizado por el zorro, observación advertida también en otros estudios similares.
La localización de restos óseos que han sido dispersados en grandes áreas es una tarea difícil, pero a veces necesaria para cualquier agencia de aplicación de la ley. Este estudio evaluó cómo el comportamiento y el patrón de dispersión para las tres especies podría ayudar a la búsqueda y recuperación de restos esqueléticos. Algunos estudios han utilizado restos de cerdo para emular las modificaciones sufridas a un cuerpo de un niño, encontrando que la actividad de los carroñeros redujo los especímenes a elementos esqueléticos para el día 5 de IPM. En el presente estudio, la utilización de un cerdo de 25kg, vestido sería compatible con un cadáver de un niño de ocho años que, bajo las circunstancias estudiadas, podría potencialmente desarticularse en un periodo de dos semanas.
Todos los carroñeros observados, mostraron diferentes comportamientos y patrones de interacción con la carroña, prefiriendo buscar en diferentes momentos del día, en diferentes etapas de descomposición y diferentes condiciones climáticas. La identificación de los carroñeros y sus comportamientos de dispersión típicos puede ayudar en la búsqueda de restos óseos, así como interpretaciones de trauma, condición y deposición de un cuerpo humano.
La intención final de esta investigación es proporcionar un marco contextual local y estacional para la interpretación de las alteraciones post depositacionales de restos biológicos por carroñeros locales. Aunque esperamos que los animales actúen sobre restos biológicos que emulan a los humanos de una manera predictiva, una comparación con la literatura muestra que los patrones generales como los que se encontraron en esta investigación son frecuentes. Sin embargo, afirmar correlaciones más específicas por el momento, son difíciles de sostener y sería necesario una revisión de las mismas para concluir la validación final de los resultados. Se pretende, además, presentar la importancia del marco ecológico evolutivo que brinde el debate teórico sobre las relaciones entre los recursos y la etología de los carnívoros en contextos peri urbanos.

