











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
El proceso desde la geminación al establecimiento de plántulas, es uno de los hechos más importantes del ciclo de vida de las plantas (Benech-Arnold y Sanchez 2004, Filho 2015). El éxito de este proceso se origina tanto en la acción de diversos factores externos, temperatura, oxígeno, luz, potencial hídrico del medio, etc., como así también en las características de las propias semillas, ya sean genéticas o derivadas de las condiciones ambientales durante su formación, madurez y conservación (Bewley 1997, Bewley y Black 1994, Dantas et al. 2015, Fhilo 2015, Shaban, 2013). Con frecuencia se ha encontrado que las semillas de diferentes poblaciones o de diferente origen geográfico varían en los requisitos de germinación y en el grado de dormición (Bevington 1986, Dorne 1981, Mc Williams et al. 1968, Pegtel 1985) en relación con la madurez de la semilla y las condiciones ambientales durante su desarrollo (Chaisurisri et al. 1992).
Si bien la humedad, el oxígeno y la temperatura son esenciales para la germinación, ciertas especies también requieren luz. La respuesta a la luz de semillas de varios cientos de especies ha sido estudiada (McDonald y Copeland 1999), encontrándose especies cuya germinación fue promovida por la luz, otras por la oscuridad, y otras resultaron indiferentes a esta condición (Baskin y Baskin 2014, Mayer y Poljakoff-Mayber 1982, McDonald y Copeland 1999). Particularmente en las especies silvestres se ha encontrado gran variabilidad de respuestas frente a este factor (Mayer y Poljakoff-Mayber 1982).
Uno de los principales factores ambientales que afecta la germinación es la temperatura (Lindow et al. 2017), y los modelos de tiempo térmico han permitido estudiar las temperaturas cardinales para la germinación (Batlla y Benech-Arnold 2015). Para especies nativas en general, la respuesta de la germinación a diferentes regímenes de temperaturas son pocos conocidas. Actualmente existe un gran interés en el estudio de estas especies dado que su cultivo presenta frecuentemente ventajas, respecto a las no nativas, tales como no requerir agua o fertilización adicional y/o un menor control de plagas y enfermedades. Por otro lado, se reduce el riesgo de la introducción de especies que puedan volverse invasoras (Torres et al. 2008). A estos aspectos, interesantes a los fines de conservación o preservación del medio ambiente, se suman los potenciales usos alimenticios, ornamentales, medicinales, entre otros (Barboza et al. 2009, D´Alfonso y Scaramuzzino 2018, Eynard et al. 2017).
Particularmente el género sudamericano Rhodophiala C. Presl (Amaryllidaceae) incluye especies herbáceas y bulbosas que tienen gran valor potencial como medicinales (Lizama-Bizama et al. 2018, Noguera Serrano et al. 2017) y algunas de ellas se cultivan como ornamentales (Hurrell y Roitman 2009). Este uso se sustenta en sus floraciones con atractivos colores, la rusticidad y la adaptabilidad a condiciones con ciertas restricciones climáticas y edafológicas. La mayoría de las especies de este género se distribuyen en Chile (Hurrell y Roitman2009). En la Argentina se encuentra representado sólo por 2 especies: R. mendocina (Phil.) Ravenna que se encuentra en Chubut, La Pampa, Mendoza, Neuquén, Río Negro y San Juan, y R. bifida (Herb.) Traub. hallada en Buenos Aires, Corrientes, Entre Ríos y Misiones y en Brasil, Paraguay y Uruguay (Zuloaga et al. 2019).
Rhodophiala bifida (azucenita del campo) es una hierba bulbosa, con hojas lineares, subcarnosas, que aparecen después de la floración. Florece a fines de verano y principios de otoño, sus vistosas flores con tépalos rojo carmín, rojo bermellón, rojo vinoso, purpúreos, rosados o blancos, dispuestos en inflorescencias con hasta siete flores (Hurrell y Roitman 2009). Estas características le confieren valor ornamental, y por esta razón se la cultiva (Dimitri 1987). Además, sus bulbos contienen alcaloides, entre ellos presenta montanina a la que se le atribuye actividad antimicrobiana y otras propiedades de interés farmacológico (Castilhos et al. 2007, Reis et al. 2019). Se reproduce por semillas que son dispersadas por el viento (Hurrell y Roitman 2009). Cada fruto (cápsula con tres lóculos) contiene numerosas semillas, aproximadamente 80 a 90 y con peso de 100 semillas relativamente alto (Echeverría y Alonso 2010). Éstas son planas, oblicuamente aladas, de color castaño oscuro a negras, brillosas, de contorno deltoide (Echeverría y Alonso 2010, Hurrell y Roitman 2009). Aunque la multiplicación por bulbos ha sido el método utilizado para la obtención de plantas para su cultivo (Dimitri 1987), en los últimos años se ha probado hacerlo por semillas (Echeverría y Alonso 2010, Hurrell y Roitman 2009). De este modo se protegen las poblaciones naturales de esta especie nativa, que al menos en la provincia de Buenos Aires, ha sido considerada vulnerable (Delucchi 2006).
La propagación agámica y sexual de algunas especies de este género, entre ellas R. bifida, han sido objeto de diversos estudios (Echeverría y Alonso 2010, Maritano et al. 2008, Noguera Serrano et al. 2017, Rosselló 2006, Rossetti et al. 2016), aunque los factores de la germinación no han sido analizados exhaustivamente. El objetivo del artículo es contribuir al conocimiento de las características fisiológicas básicas de la germinación y perduración de semillas de Rhodophiala bifida.
MATERIALES y MÉTODOS
Material estudiado y unidad experimental: Las semillas de Rhodophiala bifida se recolectaron durante marzo de 2017 y abril de 2018 en un campo colindante a la Facultad de Agronomía de la Universidad Nacional del Centro de la provincia de Buenos Aires (S 36° 45´ 990” WO 059° 52´ 971”). Este sitio está ubicado en Azul, provincia de Buenos Aires, dentro de la región denominada Pampa Deprimida. Ha sido caracterizado como una planicie deprimida con geoformas de relieve positivo (lomas), con presencia de tosca calcárea a diferentes profundidades del perfil del suelo; el tipo de vegetación dominante es el flechillar (Valicenti y Cachenaut 2018). Los ejemplares de R. bifida fueron identificados (Fabris 1969, Hurrell y Roitman 2009) y depositados en el herbario FAA (Facultad de Agronomía de Azul) (Thiers 2020) según los registros 9515 y 10020.
Luego de la recolección de las semillas, se completó el secado distribuyendo las cápsulas en una mesada sobre papel madera, trillándolas una vez abiertas y dejándolas al aire por una semana. Para las semillas a evaluar su germinación luego de un año de su producción, ante la falta de información publicada sobre las condiciones y naturaleza específica, se optó conservarlas durante ese período a una temperatura no fluctuante y reproducible (heladera a 10 °C) con la humedad alcanzada en las condiciones estacionales y en tubos polistores de 1,3 cm x 3,5 cm y dentro de doble bolsa de polietileno transparente.
Los ensayos de germinación se realizaron en cajas de Petri plásticas en las que se colocaron 50 semillas sin hidratar ni desinfectar previamente, descartándose aquellas semillas de consistencia papiráceas como evidencia de su condición de semilla vacía. La siembra se realizó sobre una hoja de papel de filtro y cuatro hojas de papel toalla, humedecidas con 4 ml de agua destilada. A los fines de evitar la desecación, cada caja fue colocada individualmente dentro de un envase plástico transparente con tapa sobre una gomaespuma impregnada en agua. Los envases plásticos luego fueron colocados de a pares en bandejas plásticas con agua y envueltas con bolsas de polietileno trasparentes, con el fin de preservar humedad próxima a la saturación y contribuir en el mantenimiento de la temperatura (Figura 1).

Figura 1 Sistema de incubación para preservar la humedad y mantener la temperatura. Se puede observar la caja de Petri, conteniendo las semillas, apoyada sobre gomaespuma embebida en agua dentro de un envase plástico con tapa. Los envases, además, se colocaron de a pares en bandejas con agua y ésta dentro de una bolsa transparente de polietileno.
Germinación a diferentes temperaturas y tratamiento luz/oscuridad: se realizó con las semillas recolectadas en 2017, iniciando la experiencia 7 días después de su recolección. Las temperaturas impuestas fueron de: 7; 14; 19; 25; 28 y 33°C (+/- 1 °C), y el fotoperíodo fue de 12 horas de luz blanca, suministrada por tubos fluorescentes. Las temperaturas y el fotoperíodo se verificaron dentro de las cámaras, estufas y heladeras de cultivo, en forma puntual en cada momento de muestreo (mediante termómetros de mercurio) y seguidos en forma horaria, durante períodos de al menos cinco días, mediante dispositivos data-logger (METEO Cavadevices). Se incluyó un tratamiento de oscuridad continua y 19 +/- 1 °C (T19*). En éste la siembra fue realizada bajo luz verde, las cajas envueltas en papel aluminio y los envases plásticos cubiertos con polietileno negro de 4 micrones para su incubación. Todos los tratamientos se realizaron con cuatro repeticiones.
Germinación de semillas frescas y de un año de edad: se realizó con semillas recolectadas en dos cosechas: 2017 y 2018. Las de 2017 se utilizaron en estado fresco, con 7 días desde la cosecha (2017a), y luego de un año de almacenamiento en heladera a 10 °C (2017b). Las semillas de 2018 se utilizaron en estado fresco, contando con poco más de dos semanas de recolectadas al momento de iniciar la prueba. El ensayo de germinación se condujo de la misma manera que los ensayos de 2017, con 4 repeticiones de 50 semillas en cajas de Petri sobre cuatro papeles toalla y uno de filtro, agregados 4 ml de agua destilada y colocadas en envases plásticos con gomaespuma y agua. La temperatura de incubación fue a 19 +/- 1 °C y con fotoperíodo de 12 horas.
Mediciones: En el ensayo a diferentes temperaturas con iluminación se realizó un seguimiento de la germinación en el tiempo. Al tratamiento de oscuridad (T19*) no se le hizo seguimiento en el tiempo y se dio por concluido cuando finalizó el tratamiento de 19 °C a luz. La frecuencia de los muestreos fue cada 3 días para los dos tratamientos con temperaturas extremas. Para el resto de los ensayos en diferentes temperaturas y el ensayo de semillas frescas, y de un año de edad de colectadas, se realizaron registros diarios hasta el inicio de la germinación y a partir de ese momento se intensificaron los registros cada 12 horas, aproximadamente. Las mediciones se dieron por finalizadas cuando la germinación se consideró total (100 % o sólo semillas con evidencias claras de putrefacción) o, alternativamente, cuando no se registró ningún caso de germinación a las 60 horas del último muestreo.
Una vez finalizado cada tratamiento se completaron los resultados de número de semillas germinadas (G) (Figura 2.A) con una inspección de aquellas que no lo hicieron. A las semillas no germinadas se las clasificó, siguiendo los criterios generales del ISTA (2006), en los siguientes estados seminales: vacías (V), muertas (M) (Figura 2.B) y firmes a la presión, diferenciándolas entre rojizas (Fr) y blancas (Fb) (Figura 2.C y D, respectivamente) según la coloración de la superficie del endosperma, observación realizada luego de ser retiradas las cubiertas seminales. Con estos datos se calcularon los porcentajes de cada estado final seminal (%G; %Fb; %Fr y %M) sobre un total que excluyó las semillas vacías. También se calculó el porcentaje de la germinación acumulada en el tiempo (%Gt).
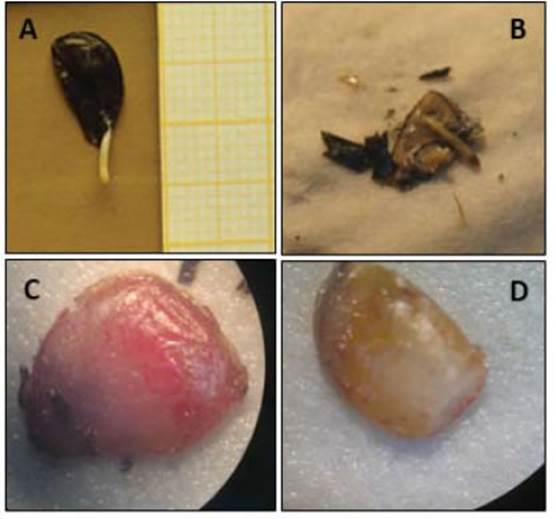
Figura 2 Aspecto de las semillas de R. bifida. Estados encontrados al finalizar los tratamientos: Semilla germinada (A); semillas muertas (B), semilla firme y roja (C) y semilla firme y blanca (D).
Manejo y Análisis de datos: los porcentajes finales de estados seminales fueron analizados mediante ANOVAS y los tratamientos comparados mediante la prueba de LSD Fisher, luego de ser normalizados apropiadamente (Sokal y Rohlf 1986) y verificada la homocedasticidad. Para cada unidad experimental los %Gt se ajustaron en función del tiempo (T) mediante la siguiente función: %Gt= A*(1-1/(1+(T/B)^k)). El significado biológico de los parámetros correspondió a: Porcentaje Máximo de Germinación (parámetro A); Tiempo Medio de Germinación (parámetro B) y el valor adimensional de la Uniformidad en la Germinación (parámetro K). Los valores de los parámetros obtenidos fueron analizados mediante ANOVA y contrastados mediante la prueba de LSD Fisher (α ≤ 0.05). A partir de los Tiempos Medios de Germinación (B) obtenidos en cada temperatura, se estimó la temperatura base (Tb) de la germinación y el Tiempo Térmico requerido para completar la germinación (TTg). Los parámetros obtenidos fueron analizados mediante un ANOVA y contrastados con la prueba LSD Fisher (α≤0.05). Todos los procedimientos estadísticos se realizaron con el programa InfoStat (Di Rienzo et al. 2019).
RESULTADOS Y DISCUSIÓN
Germinación a diferentes temperaturas y tratamiento luz/oscuridad
En ninguna de las temperaturas empleadas se evidenciaron problemas sanitarios destacables, registrándose al final de la experiencia porcentajes de germinación (%G) cercanos o superiores al 90 %, salvo a 33 °C, temperatura en la que fue un 10 % menor (Tabla 1). Los mayores valores de %G se registraron a 19°C tanto en luz como en oscuridad (Tabla 1). El porcentaje relativamente alto de semillas muertas (%M) a 33 °C explicó, en parte, la menor germinación antes comentada (Tabla 1). Al comparar los resultados con los obtenidos a 20°C por Rossetti, Rojahn y Costa (2016), bajo alternancia de temperaturas por Echeverría y Alonso (2010) y a 25 ºC por Rodrigo et al. (2006), se observan valores similares y coincidentes, que permiten evidenciar que esta especie, en una proporción elevada de semillas y en un amplio rango de temperaturas, no presenta dificultades para la germinación, aspecto concomitante con la ausencia de dormición.
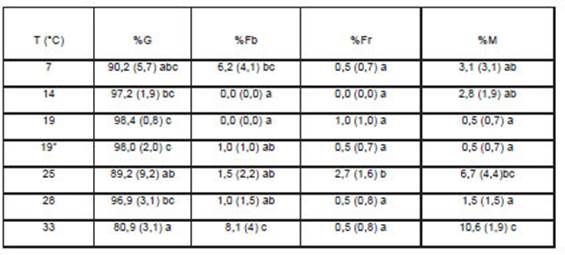
Tabla 1: Promedios del porcentaje de cada estado seminal obtenidos a diferentes temperaturas. Los promedios corresponden a cuatro repeticiones y entre paréntesis se especifica el desvío estándar. Letras diferentes dentro de una columna indican diferencias significativas (α≤0.05). %G= porcentaje de semillas germinadas; %Fb= porcentaje de semillas firmes y blancas; %Fr= porcentaje de semillas firmes y rosadas; y %M= porcentaje de semillas muertas
Al final del ensayo, el porcentaje de semillas aparentemente sanas, firmes y de coloración blanca (%Fb) luego de su transformación conveniente para el análisis estadístico, presentó distribución normal pero no homogeneidad de varianza. Sin embargo, la falta de un patrón visible entre los residuos y los valores predichos de esta variable alentó a continuar con el análisis de la varianza, aunque sus resultados deben ser tomados con precaución. A 33 °C se obtuvo el valor promedio más alto (8,1 %) de semillas Fb, encontrándose un valor similar sólo a 7 °C (Tabla 1). Estos valores contribuyeron, en parte, a explicar el menor porcentaje de semillas germinadas en esas temperaturas, particularmente en 7 °C. En las temperaturas de 14 y 19 °C (luz) no se hallaron semillas Fb (α ≤ 0.05, Tabla 1). Por otro lado, el porcentaje de semillas firmes de coloración roja-rosadas (Fr) no presentó efecto de la temperatura y sus valores fueron relativamente bajos (Tabla 1). Estas semillas presentaron el endosperma con aspecto granuloso transparente. Tanto las características de coloración y aspecto podrían corresponder a semillas con irregularidades en el grosor, que se observaron en el momento de la siembra pero que no fueron convenientemente marcadas para su seguimiento. Por tanto, es necesario realizar otro tipo de estudio para determinar si estos estados seminales corresponden a defectos genéticos y/o de desarrollo seminal o, alternativamente a semillas capaces de germinar dentro de ciertas condiciones y dar plántulas normales.
En cuanto al porcentaje de semillas muertas (%M) el mayor valor se obtuvo en la mayor temperatura empleada (33 °C) alcanzando al 10% de las semillas (Tabla 1). Si bien esta mortalidad no es muy elevada, resulta contrastante con la termoinhibición presentada a 33 °C por Habranthus tubispathus (Manfreda et al. 2017), especie nativa de la misma familia con la que comparte ambientes y otras características del desarrollo, como floraciones coincidentes a fines de verano. En cuanto a la termoinhibición, ha sido propuesta como una adaptación de plantas anuales de invierno a climas con inviernos húmedos y veranos secos (Derakhshan et al. 2018), adaptación que impide que la germinación ocurra mientras persisten las temperaturas estivales. R. bifida además de carecer de esta adaptación, mostró ser perjudicada por la alta temperatura.
La germinación conducida en oscuridad a 19 °C (19*, Tabla 1) mostró la ausencia de requerimiento de luz para la germinación, obteniéndose altos porcentajes de germinación y similares porcentajes de semilla muertas, firmes y rojas, a los obtenidos a 19 °C con luz (α ≤ 0,05; Tabla 1). Esta característica de fotoblastismo neutro fue coincidente con lo reportado por Echeverría y Alonso (2010) en semillas de R. bifida provenientes de Gral. Pueyrredón (Sierra de los Padres, Provincia de Buenos Aires).
El ajuste a función de la germinación acumulada para cada unidad experimental fue descrito adecuadamente por la ecuación utilizada, con menor o igual SC Error que la función de Gompertz (datos no mostrados), función frecuentemente utilizada en los estudios de dinámica de la germinación.
Los porcentajes de germinación final obtenidos por esta ecuación, en el rango entre 7 y 25 °C, no evidenciaron diferencias significativas (α ≤0,05; (Tabla 2) y tomaron valores próximos a los de su medición directa (Tabla 1). Dentro del mismo rango térmico los Tiempos Medios de Germinación estimados (parámetro B de la ecuación) presentaron efectos significativos en respuesta a la temperatura (Tabla 2), mostrando una relación inversamente proporcional con el aumento de temperatura. Se registró el mayor tiempo medio a 7 °C y el menor a 25 °C (α ≤0,05; Tabla 2). En cuanto al parámetro K de la ecuación, dentro de este rango térmico, no mostró diferencias significativas (α ≤0,05; Tabla 2). Este parámetro indica cuán extendida es en el tiempo la germinación, por lo tanto cuanto más homogénea y más concentrada es la germinación, mayor es el valor de K.
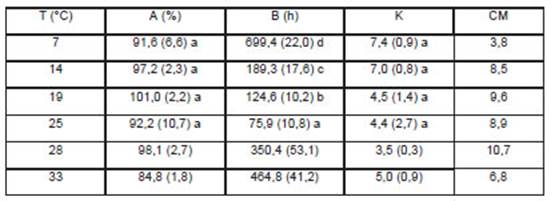
Tabla 2 Promedios de los parámetros de las curvas de germinación de R bifida para cada temperatura. Los desvíos estándar qué están entre paréntesis. Letras diferentes dentro de una columna indican diferencias significativas (α ≤ 0.05). A=Porcentaje Máximo de Germinación (%); B=Tiempo Medio de Germinación (h); K=Uniformidad en la Germinación; CM= Cuadrado Medio del Error.
El parámetro B es de particular significancia para evaluar las características del desarrollo de una especie frente a la temperatura, ya que la inversa de este parámetro es un estimador de la velocidad de desarrollo. Se observó que la velocidad de germinación (Vg) en respuesta a la temperatura no se apartó del modelo general de tiempo térmico (Línea punteada Azul, Figura 3) con una To que se ubicó alrededor de los 25 °C (Figura 3). Por encima de esta temperatura la Vg disminuyó abruptamente en 28 °C (Figura 3). La estimación de la Tb y del TTg, obtenidos por el ajuste lineal de la fase positiva de la respuesta (entre 7 y 25 °C, Figura 3), fueron aproximadamente de 4,83 °C y 1666 °Ch respectivamente.
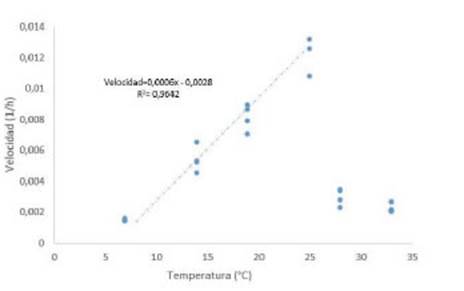
Figure 3 Germination Speed obtained in R. bifida at different temperatures. The blue dotted line corresponds to represents lineal fit between Speed and Temperature (°C) for the temperature range with positive relationship. Equation and Person coefficient (R2) obtained are showed.
Los tres parámetros de la ecuación para 28 y 33 °C fueron excluidos del análisis estadístico anterior, ya que sus valores presentaron alta variabilidad, no presentaron homogeneidad de varianza ni distribución normal en el conjunto de los datos anteriores. Al hacer el análisis comparativo de los valores promedios de estos parámetros y sus desvíos (Tabla 2) se pudo observar que el valor de A fue menor en 33 °C que en 28 °C, mientras que los parámetros B y K fueron mayores. Esto último indica que, la temperatura aumentó la cantidad de horas para completar el 50 % de germinación y, por lo tanto, disminuyó la Vg, acompañándose con un aumento de la uniformidad en la germinación.
Germinación de semillas frescas y de un año de edad
Todas las semillas cosechadas, tanto las frescas de 2017 (2017a) y de 2018 como las almacenadas por un año (2017b), alcanzaron valores superiores al 98 % de germinación (Tabla 3), sin mostrar efecto del año de la cosecha ni del año de almacenamiento (α ≤0,05). La inspección de las semillas que no germinaron se correspondió fundamentalmente a su mortalidad (11 % y Desvstd = 6, dato no mostrado). Estos altos valores de germinación de semillas recientes y de un año de edad no son coincidentes con los registrados por Maritano et al. (2008), quienes reportaron para R. bifida valores máximos de 85 % en semillas frescas y del 65 % en semillas de un año. La falta de coincidencia puede deberse tanto a diferencias poblacionales, condiciones de almacenamiento y/o al empleo de técnicas de desinfección. Estas últimas pueden ocasionar daño y/o deterioro de las semillas y afectar negativamente el porcentaje final de la germinación (Abdul-Baki 1974, Baskin y Baskin 2014, Ponzio 1998). Este aspecto fue identificado por Noguera Serrato et al (2017) en R. mendocina donde evidenciaron mayores porcentajes de germinación en semilla sin desinfectar, embebidas y en oscuridad.
Tabla 3: Promedios de los porcentajes de germinación (%G) y de los parámetros de la curva de ajuste (A, B y K, ver leyenda de Tabla 2 y Materiales y Métodos) para semillas frescas (2017a y 2018) y luego de un año de almacenaje a 10°C (2017b). Los desvíos estándar se presentan entre paréntesis. Letras diferentes dentro de una columna indican diferencias significativas (α ≤ 0.05).
Table 3: Mean values of gemination percentages (%G) and average of parameters of curves (A, B and K, shown in table 2 and Material and Methods) for fresh seeds (2017a and 2018) and after one year of storage at 10°C (2017b). The standard deviation is shown in parentheses. Different letter within a column indicate significant differences (α ≤ 0.05).

En cuanto a los ajustes obtenidos a la curva de germinación, los parámetros A y K no presentaron efecto de la edad ni del año de cosecha (Tabla 3). El parámetro B mostró, por un lado, que el almacenamiento por un año redujo significativamente en 20 h el tiempo medio de germinación (α ≤ 0,05; Tabla 3). Por otro lado, mostró diferencias significativas según el año de cosecha, siendo menor en 2018 que en 2017 (α ≤0,05; (Tabla 3). Mientras que la primera diferencia puede relacionarse con una posible posmaduración de las semillas, la obtenida entre diferentes años de cosechas es posible atribuirlas a efectos maternos. Ya que son múltiples las condiciones ambientales durante la formación de las semillas que pueden afectar la germinación, tales como la nutrición parental (Fenner 1986, Marshall 1986, Parrish y Bazzaz 1985, Stratton 1989), el nivel hormonal (Gutterman 1980), el estrés por sequía (Sawhney y Naylor 1980), la longitud del día y la calidad de la luz (Gutterman 1980), y la temperatura (Alexander y Wulff 1985) no fue posible identificar uno en particular que justifique las diferencias encontradas en el Tiempo Medio para las semillas en estudio.
CONCLUSIONES
Las semillas de R. bifida evidenciaron porcentajes de germinación alrededor del 90% en temperaturas de 7, 14, 19, 25 y 28 °C. Los mayores porcentajes se obtuvieron a 19 °C, tanto en luz como en oscuridad. El menor valor de %G se obtuvo a 33 ºC, y coincidió, por un lado con el aumento de la mortalidad y, por otro lado, con un mayor porcentaje de semillas firmes y blancas.
La dinámica de germinación de R. bifida, en todos los tratamientos, pudo ser descripta adecuadamente con la función utilizada, dando base uno de sus tres parámetros a la estimación de la velocidad de germinación. Esta última respondió linealmente a la temperatura en el rango de 7 y 25 °C, disminuyendo abruptamente a 28 y 33 °C. En base a esta respuesta lineal se estimó la Tb y el TTg, siendo de 4,83 °C y 1666 °Ch respectivamente.
Luego de un año de almacenamiento las semillas de R. bifida no presentaron dificultad en la germinación, y se obtuvieron valores cercanos al 100 %. Sin embargo, el Tiempo Medio de Germinación disminuyó en 20 h respecto a las recientemente cosechadas, siendo también afectado significativamente por el año de cosecha, por lo que se evidencia una modificación de su calidad fisiológica y no se descarta un efecto materno. Los altos porcentajes de germinación en un amplio rango de temperaturas, la ausencia de problemas sanitarios, de dormición y de termoinhibición en las temperaturas superiores, el fotoblastismo neutro, la buena longevidad fueron las principales características de la fisiología de la germinación de las semillas de R. bifida evidenciadas por este trabajo.

