











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Prosopis alba Griseb. es una especie nativa de Sudamérica, con gran importancia en la composición arbórea de zonas áridas y semiáridas del mundo. Es una especie de gran potencial para la producción de material forestal y la de mayor importancia económica dentro del género. P. alba es empleada tradicionalmente de forma intensiva como madera para muebles, en la construcción, para la producción de leña y carbón, en la alimentación animal dentro de sistemas silvopastoriles y en la alimentación humana. Por otra parte, es una especie importante como fijadora de nitrógeno (Felker et al. 2001, Ewens y Felker 2010).
La propagación vegetativa de especies arbóreas de importancia forestal constituye una herramienta valiosa desde el punto de vista productivo ya que permite acelerar el desarrollo de la forestación reduciendo los turnos de corte y la propagación de materiales más productivos, de mejor calidad y con características silvícolas de importancia. A pesar del elevado interés por realizar la producción clonal a gran escala de P. alba, hasta la actualidad no se han logrado protocolos eficientes que garanticen el establecimiento, desarrollo y regeneración de esta especie (Felker et al. 2005, Felker et al. 2008, Araujo Vieira de Souza et al. 2014).
La técnica de propagación vegetativa a través de estacas asociada a la aplicación de ácido indolbutírico (IBA) puede ser utilizada para la propagación de especies forestales nativas de interés comercial. Su utilización puede ser limitada por la falta de métodos eficientes de rejuvenecimiento de material adulto y de técnicas de manejo adecuadas en el ambiente de propagación (Dias et al, 2015).
En P. alba la regeneración mediante estacas resulta muy difícil de emplear dada la baja capacidad de enraizamiento de esta especie (Ewens et al. 2012, Araujo Vieira de Souza et al. 2014). En algunas plantas, la capacidad de enraizamiento de las estacas se relaciona con su estructura anatómica y con su grado de lignificación. La reducción de la lignificación de los tejidos puede aumentar la capacidad de enraizamiento en estacas y miniestacas de especies leñosas (Biasi et al. 2002, Borges et al. 2011, Schwarz et al. 2015).
Como en muchos otros representantes de la subfamilia Mimosoideae, los tallos jóvenes de P. alba presentan un anillo de esclerénquima lindante con las capas más internas del córtex. Este anillo puede ser continuo o aparecer interrumpido, y puede constituir una barrera mecánica para el desarrollo de raíces adventicias (Caro et al. 2002, Evert 2006, Peixe et al. 2007, Amissah et al. 2008, Schwarz et al. 2015).
En la propagación vegetativa la utilización de las técnicas de miniestacas y de etiolación permiten obtener propágulos con un menor grado de lignificación. La técnica de miniestacas, consiste en la utilización de brotes de plantas madres, los que son rejuvenecidos a través de la ruptura de la dominancia apical por la poda drástica de plantas madres. Mientras que la etiolación consiste en la reducción de la lignificación de los tejidos a través de la reducción de la intensidad de luz a la que crecen los brotes de las plantas madres que serán utilizados como propágulos (Biasi et al. 2002, Alfenas et al. 2009, Borges et al. 2011, Araujo Vieira de Souza et al. 2014). Las técnicas de estacas y miniestacas se diferencian en función del manejo de sus plantas madres y no del tamaño de sus propágulos (Alfenas et al. 2009).
Este trabajo tuvo como objetivo observar por medio de estudios anatómicos si efectivamente ocurre una reducción del grado de lignificación de los propágulos de P. alba al utilizar la técnica de miniestacas en relación a la técnica de estacas y al aplicar la técnica de etiolación. También, se pretende evaluar la influencia del grado de lignificación en los ensayos de enraizamiento.
MATERIALES Y MÉTODOS
Los ensayos de enraizamiento de estacas y miniestacas fueron realizados en los Laboratorios de Producción Vegetal y de Morfología Vegetal de la Facultad de Ciencias Agrarias de la Universidad Nacional del Litoral (Latitud 31º 26´31´´ S y Longitud 60º56´25´´ O) en dos años consecutivos. En el momento de inducción del enraizamiento del “año 1” las plantas madres del jardín y minijardín clonal (donantes de propágulos para el enraizamiento) poseían 18 meses y en el “año 2” poseían 30 meses.
Para la preparación de las estacas fueron utilizados brotes jóvenes provenientes de un jardín clonal y para la preparación de las miniestacas fueron utilizados brotes rejuvenecidos de un minijardín clonal. El rejuvenecimiento de los brotes del minijardín se obtuvo a través de la ruptura de la dominancia apical por la poda drástica de las plantas madres.
En el ensayo de enraizamiento del “año 2” las plantas madres del minijardín clonal fueron sometidas a la aplicación de técnicas de etiolación por medio del incremento de la densidad de plantas y sombreado.
Material Vegetal
Las plantas madres del jardín y del minijardín fueron originadas de plantas provenientes del rodal semillero de la localidad de Vera (Santa Fe, Argentina) (Latitud 29º 27´55´´ S y Longitud 60º12´58´´ O). Cada planta madre representa un genotipo de medio hermanos. Lo anterior posibilita la clonación de familias superiores.
Formación y manejo del jardín y del minijardín clonal
El jardín clonal fue implantado a campo y estaba compuesto por 40 plantas madres distribuidas aleatoriamente en 4 parcelas (con 10 plantas madres cada una), con una distribución espacial de 3 x 3 m y fueron fertilizadas con 3 ml de solución Hoagland al 50% (Hoagland y Arnon 1950) cada 15 días y previo a las colectas de los brotes. Los brotes jóvenes fueron cosechados para el ensayo de enraizamiento de estacas del “año 1” y “año 2” presentando un total 108 brotes por año. Se utilizaron entre cuatro y cinco brotes por planta madre del jardín clonal en cada año.
El minijardín clonal estuvo constituido por 55 plantas madres distribuidas en 5 parcelas, con 11 plantas cada una, dispuestas al azar y mantenidas en invernadero. Para su formación fue utilizada la metodología descripta por Araujo Vieira de Souza et al. (2009) con modificaciones en el régimen de temperatura, luminosidad, riego y fertilización. El mismo fue implantado en cámara de crecimiento con fotoperiodo de 14 h, intensidad de radiación de 400 mE PAR, temperatura día/noche de 26/19 ºC y humedad relativa del 60% donde permanecieron 24 meses. Luego, fueron trasladadas al invernadero donde se sometieron a temperaturas día/noche de 26/15 ºC y humedad relativa del ambiente de 71,5%. En la producción comercial de plantines por miniestacas las plantas madres normalmente quedan de dos a cinco años en estas condiciones. Se modificó el ambiente del minijardín clonal para aplicar las técnicas de etiolación.
Las plantas madres del minijardín fueron cultivadas en macetas de 276 cm3 con sustrato de compost de residuos sólidos urbanos con 1/4 de arena (Humedad: 45%; pH: 7,1; N total: 0,9%; P2O5: 0,9 %; K2O: 0,6%). Estas fueron fertilizadas con 3 ml de solución Hoagland al 50% (Hoagland y Arnon 1950) cada 15 días y previa a las colectas de los brotes.
Para promover el rejuvenecimiento de las plantas madres y obtener los brotes rejuvenecidos con menor grado de lignificación en el ensayo de enraizamiento de miniestacas, se realizó el corte del ápice de las mismas aproximadamente a 20 cm de la base del tallo cuando éstas presentaron una altura superior a 25 cm. Los brotes rejuvenecidos para el ensayo de enraizamiento fueron cosechados de miniestacas del “año 1” y “año 2”, obteniendo180 brotes en cada año. De cada planta madre se cosecharon para los experimentos 4 brotes por año.
Ensayo de estacas del “año 2” sometido a técnica de etiolación
En el “año 2” los brotes del jardín clonal a utilizarse para el enraizamiento fueron sometidos a sombreado total a lo largo del brote a partir del momento que las longitudes de los mismos fueron de 10 cm. Para ello, se empleó plástico negro abrochado a lo largo del brote desde de su base hasta la región inferior al ápice, que permaneció sin sombreado.
Ensayo de miniestacas del “año 2” sometido a técnicas de etiolación
Las plantas madres del minijardín clonal en el “año 2” fueron sometidas a tres diferentes técnicas de etiolación con la finalidad de promover un menor grado de lignificación en el material a ser propagado.
Se incrementó la densidad de plantas del “año 2” en relación al “año 1”. La densidad de plantas madres en los ensayos del “año 1” fue de 40 plantas m-2 y en el “año 2” de 256 plantas m-2, con menor espaciamiento entre plantas madres y mayor etiolación de brotes.
En el “año 2” los brotes fueron sometidos también a sombreado parcial y total a lo largo del brote. El sombreado parcial de las plantas madres fue conferido por el uso de media sombra (70%) a partir del momento que el promedio de longitud de los brotes fue de 10 cm y la producción de brotes por planta madre igual a 3,3. El sombreado total del brote fue conferido por el uso de plástico negro abrochado a lo largo de este desde de su base hasta la región inferior al ápice.
Preparación y evaluación de estacas y miniestacas: ensayo de enraizamiento
Para la preparación de las estacas y miniestacas se empleó el procedimiento descripto por Araujo Vieira de Souza et al. (2009). Se obtuvieron brotes juveniles del jardín clonal y rejuvenecidos del minijardín clonal para inducir el enraizamiento. Para ello, se utilizó la porción apical del brote (6 cm de largo) con al menos 3 entrenudos y 3 hojas de las cuales su superficie foliar se redujo al 50%. Posteriormente, fueron sometidas a la aplicación de fungicida (2,5 cm3.L-1 de carbendazim).
Las estacas fueron sometidas a tres tratamientos con IBA solubilizado en hidróxido de potasio (KOH 1 mol. L-1), aplicado por vía líquida en la base de cada estaca durante 15 segundos a las concentraciones de 0, 3.000 y 6.000 mg.L-1. Igual procedimiento se empleó en las miniestacas, aunque las concentraciones fueron de 0, 3.000, 4.500, 6.000 y 7.500 mg.L-1.
Posteriormente, se realizó el estaqueado de las estacas y miniestacas en tubetes plásticos de 110 cm3 conteniendo sustrato Dynamics 2®. Luego fueron transferidos a cámara de enraizamiento con nebulización intermitente, humedad superior a 80%, temperatura media de 23,6 °C, temperatura media máxima de 35°C, radiación solar 20,48 mJ.m-2.día y heliofanía efectiva promedio de 9,32 h.día-1. La fertilización fue quincenal con 2 ml tubete-1 de solución nutritiva Bolle Jones (Chaves et al., 2006).
Los experimentos de enraizamiento fueron dispuestos en un diseño de cuatro bloques aleatorizados con cinco tratamientos de IBA en miniestacas y tres en estacas. Hubo nueve propágulos (miniestacas o estacas) por unidad experimental. Los ensayos fueron realizados en dos años consecutivos en primavera.
Luego de 40 días en cámara de enraizamiento se determinó la supervivencia y el porcentaje de miniestacas y estacas enraizadas.
En el material enraizado (plantines clonales) se registró altura, diámetro del tallo, peso de materia fresca de la parte aérea, número de hojas, número de foliólulos, peso de materia fresca, número y longitud total de raíces. Las raíces fueron cuantificadas mediante digitalización de imágenes utilizando el software ImageJ® (Rasband, 1997-2019).
Se utilizó el test de Lilliefors y el de Cochran para determinar la normalidad y homogeneidad de variancias, respectivamente. Posteriormente, los resultados obtenidos se sometieron a análisis de la variancia. Los datos supervivencia y enraizamiento fueron transformados en arco seno √X/100, los datos número de hojas, de foliólulos y de raíces fueron transformados por √(X+1) y los datos masa fresca y longitud de raíces fueron transformados por log10 (X+1) de acuerdo a Zimmermann (2004). Las diferencias fueron sometidas a ajustes de regresión y en el caso de ajustes cuadráticos se determinó el punto de inflexión de las curvas por derivación, posibilitando encontrar el punto óptimo de la dosis de IBA. Para la realización de los procedimientos estadísticos fueron utilizados los programas estadísticos InfoStat (Di Rienzo y col., 2010) y ASSISTAT Software (Silva y Azevedo, 2009).
Estudio de las características anatómicas
Los estudios anatómicos fueron realizados en el laboratorio de Anatomía Vegetal de la FCA-UNL. Se tomaron muestras de miniestacas y estacas de todos los tratamientos, previo a la inducción del enraizamiento y al finalizar el ensayo. Las muestras fueron tomadas a 0,5 cm de la base de las miniestacas y estacas. Luego, la mitad de estas muestras fueron fijadas en FAA (formol: ácido acético: etanol 70%, 10: 5: 85, v/v/v), y la otra mitad de las muestras fueron conservadas en freezer a -20 °C.
Las muestras fijadas en FAA fueron deshidratadas en una serie de alcohol etílico de concentración creciente, clarificadas con xilol e incluidas en parafina (Ruzin 1999). Luego, mediante micrótomo rotativo se realizaron cortes transversales seriados de 12-15 µm de espesor, los que fueron coloreados con safranina-fast green y montados en bálsamo de Canadá (D´Ambrogio de Argüeso 1986).
De las muestras conservadas a -20 °C se obtuvieron cortes transversales seriados de 10 µm de espesor, mediante micrótomo de congelación, los cuales fueron clarificados con solución de hipoclorito de sodio, lavados con agua y montados en fluoroglucinol-ácido clorhídrico para observar el grado de lignificación de los tejidos de miniestacas (sometidas y no sometidas a etiolación) y estacas (Kraus y Arduin 1997).
Además, fueron cosechados de las plantas madres brotes similares a los que fueron utilizados para la preparación de las miniestacas y estacas y de los cuales se tomaron muestras de la región apical, proximal, media y basal. Éstas fueron fijadas en FAA, deshidratadas en una serie creciente de alcohol etílico, incluidas en parafina, cortadas con micrótomo de rotación (cortes transversales seriados) y procesadas como se indicó anteriormente.
Se analizaron en todos los tejidos observados, su grado de lignificación, el desarrollo y evolución del anillo de esclerénquima a lo largo del eje y se compararon con los resultados de los experimentos de enraizamiento.
RESULTADOS
En el ensayo de primavera del “año 1” con utilización de brotes no etiolados se observó una baja supervivencia (20%) y enraizamiento (12,5%) de las miniestacas. La rizogénesis se produjo en la región del nudo o cercano al corte realizado durante su preparación. No se observó enraizamiento de las estacas, habiéndose registrado sólo un 8% de supervivencia de las mismas (Tabla 1).
En el ensayo de miniestacas de primavera del “año 2” con utilización de brotes etiolados se registraron elevados valores de supervivencia y enraizamiento (96 y 99,6%, respectivamente). Para las estacas del jardín clonal se obtuvieron mejores valores de supervivencia (20%) y enraizamiento (7,6) que en el “año 1”, pero los mismos fueron inferiores a los registrados en miniestacas (Tabla 1).
No se observaron diferencias en función de las concentraciones de IBA en el porcentaje de enraizamiento de las miniestacas en el ensayo de primavera del “año 2”. Para las concentraciones de IBA de 0, 3.000, 4.500 y 6.000 mg. L-1 se obtuvo un 100% de enraizamiento y con la dosis de 7.500 mg. L-1 fue del 98% (Tabla 2). No hubo correlación entre el grado de lignificación del anillo de esclerénquima y la concentración de IBA aplicado.

Tabla 1 Porcentajes de supervivencia (SUPERV) y enraizamiento (ENR) en los ensayos de miniestacas y estacas en los años 1 y 2.
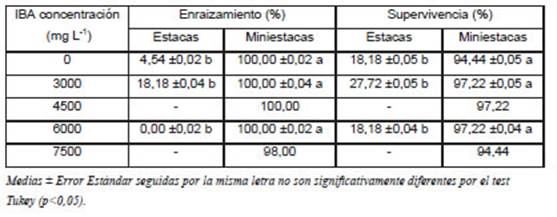
Tabla 2 Porcentaje de enraizamiento y supervivencia de estacas y miniestacas de Prosopis alba sometidas a etiolación y tratadas con diferentes concentraciones de ácido indolbutírico (IBA).
La utilización de IBA influenció el número de hojas y foliólulos, número, peso de materia fresca y longitud total de raíces. Se observó mejor desarrollo radical hasta la dosis de 6.000 mg. L-1, mientras que dosis superiores tuvieron efecto negativo (Fig.1). El número de hojas de los plantines se incrementó hasta un punto óptimo de 4.306,7 mg L-1 (valor obtenido por derivación del punto de inflexión de las curvas generadas por el análisis de regresión de ajustes cuadráticos), registrándose una disminución del número de hojas a partir de esta concentración. Igual respuesta se observó en relación a la cantidad de foliólulos hasta su punto óptimo 4.113,5 mg L-1.
La cantidad, peso de materia fresca y longitud de raíces de los plantines tuvieron los siguientes puntos óptimos: 4.840, 3.480 y 3.726 mg L-1 (Fig.1).
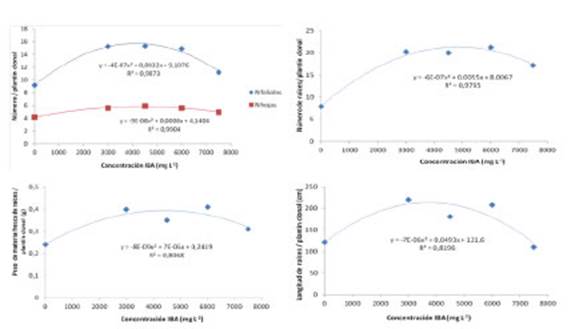
Fig. 1 Efecto de la concentración de ácido indolbutírico (AIB) sobre variables de la parte aérea y radical de los plantines clonales de P. alba obtenidos a través de miniestacas y sometidas a etiolación a los 40 días.
En los cortes transversales de la región apical de brotes rejuvenecidos de plantas madres del minijardín clonal se observó una epidermis uniestratificada, una capa subepidérmica de colénquima, pocas capas de clorénquima, algunas células taníferas, una capa cortical interna de células grandes (vaina amilífera), el anillo de esclerénquima en formación, haces vasculares colaterales y una región medular con células parenquimáticas grandes en comparación con el tamaño de las células del córtex (Fig.2, A y B).
Desde el ápice hacia la base del brote fue aumentando el número de haces vasculares en formación, apareciendo diferenciados, en primer lugar, elementos de floema y luego elementos de xilema y a un mismo nivel se observaron haces vasculares en distintos estados de diferenciación (Fig. 2). También hacia la región proximal del brote, se observó la aparición de cambium vascular, que resultó temprana en los haces vasculares más grandes, y fue más retrasada en los medianos y pequeños; presentándose áreas interfasciculares sin cambium vascular notable (Fig. 2). En paralelo con la diferenciación de cada haz vascular se fue diferenciando un grueso casquete floemático de esclerénquima. Las células más externas del mismo fueron las que primero engrosaron sus paredes; mientras que las células más internas persistieron mayor tiempo como células de paredes delgadas (Fig. 2, C y D). Finalmente, en regiones más basales del brote todo el casquete floemático de fibras presentó paredes celulares gruesas y esclerificadas (Fig. 2, F). En general los casquetes de esclerénquima fueron aumentando de tamaño proporcional al tamaño que fue tomando el haz vascular. Las células que formarían, en regiones más diferenciadas, el anillo de esclerénquima, mostraron en las regiones apicales una estructura constituida por casquetes floemáticos unidos por regiones de menor cantidad de estratos celulares (Fig. 2).
Con el incremento del crecimiento secundario se observó la típica estructura de las especies leñosas, en las cuales el cambium vascular se dispone entre anillos de xilema secundario hacia el interior del órgano, y de floema secundario hacia el exterior (Fig. 2, E y F). Se determinó que el xilema secundario en las regiones fasciculares era de estructura diferente al xilema secundario de las regiones interfasciculares. Estas últimas, presentaron menor proporción de vasos y se diferenciaron radios vasculares (Fig. 2). Las células de los radios xilemáticos esclerificaron sus paredes, en tanto que los radios floemáticos permanecieron parenquimáticos y se fueron ensanchando radialmente hacia la periferia (Fig. 2). Por fuera del floema secundario apareció el anillo de esclerénquima formado por los casquetes de esclerénquima y regiones entre dichos casquetes (Fig. 2), los que estaban formados por varias capas de fibras de pequeño diámetro y gruesas paredes fuertemente lignificadas. En las regiones entre casquetes se observó 1-2 capas de células grandes de paredes más delgadas y menos lignificadas, siendo algunas células no lignificadas (Fig. 3, A y B). Las regiones entre casquetes se asociaron a regiones lindantes con las áreas interfasciculares y con la parte distal de los radios floemáticos. Estas regiones aparecieron como posibles discontinuidades en el anillo de esclerénquima en material menos lignificado. El espesor y el grado de lignificación de las paredes celulares de la región entre los casquetes de esclerénquima, aumentó hacia la base de la miniestaca (Fig. 2).

Fig. 2 Brotes rejuvenecidos de plantas madres del minijardín clonal de Prosopis alba de las cuales se extrajeron propágulos para preparación de miniestacas. A y B: corte transversal de entrenudo de la región apical; C y D: región media del brote; E y F: región basal del brote. Referencias: casquete de esclerénquima (ce); cambium vascular (cv); floema (fl); periciclo (p); región entre casquetes de esclerénquima (rec); xilema (xl).
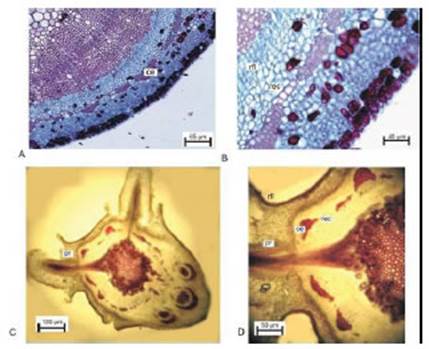
Fig. 3 Miniestacas de Prosopis alba año 1. A: corte transversal de entrenudo de miniestaca antes de iniciar los ensayos de enraizamiento; B: detalle ampliado de A; C: miniestaca enraizada; D: detalle ampliado de C. Referencias: casquete de esclerénquima (ce); región entre casquetes de esclerénquima (rec); radio floemático (rfl); primordio radical (pr).
El grado de lignificación de los casquetes de esclerénquima y de las regiones entre casquetes fue mayor en las estacas respecto de las miniestacas (Fig. 4). Del mismo modo dicho grado de lignificación fue mayor en las miniestacas y estacas del primer año, respecto a las del segundo año (Fig. 4, C - E).
En estacas y miniestacas de P. alba en las que se logró el desarrollo de raíces adventicias se observó que los primordios radicales atravesaban el anillo de esclerénquima compuesto a través de las regiones entre casquetes de esclerénquima (Fig. 3, C y D). En las miniestacas se observó enraizamiento en las regiones que mostraron menor grado de lignificación en las paredes celulares, es decir en las regiones entre casquetes de esclerénquima. Para las mismas condiciones de enraizamiento, miniestacas de P. alba que presentaron mayor grado de lignificación en esas regiones, no enraizaron (Fig. 4, D).
El menor grado de lignificación de los tejidos promovió tasas superiores de enraizamiento de estacas y miniestacas en P. alba.

Fig. 4 Variaciones en los grados de lignificación y discontinuidad de los anillos esclerenquimáticos de miniestacas y estacas de Prosopis alba. A: miniestaca del año 1 que no enraizó; B: estaca del año 1, con mayor grado de lignificación, que no enraizó; C: miniestaca del año 1, con menor grado de lignificación, que enraizó y que presentó mayor producción de raíces a los 40 días; D: miniestaca del año 1 con mayor grado de lignificación, que enraizó pero que presentó baja producción de raíces a los 40 días; E: grado de lignificación de miniestaca del año 2, crecida en mayor condición de etiolación y que presentó 99,6 % de enraizamiento.
DISCUSIÓN
Las diferencias en la anatomía de las estacas y miniestacas de P. alba se relacionaron con variaciones en el grado de lignificación del anillo de esclerénquima. El porcentaje de enraizamiento logrado estuvo afectado directamente por el grado de lignificación del propágulo utilizado; cuanto menor fue la lignificación de los brotes utilizados para propagación, mayores fueron las tasas de enraizamiento.
Miniestacas etioladas presentaron menor grado de lignificación que estacas y que miniestacas no etioladas y, consecuentemente, lograron mayores porcentajes de enraizamiento, número y longitud total de raíces. La formación de raíces adventicias es una etapa necesaria en la propagación vegetativa, y debido a ello el interés de este proceso en la silvicultura (Oinam et al. 2011). La incapacidad de desarrollar raíces adventicias desde estacas o la pérdida de dicha capacidad que ocurre con la edad de la planta ocasionan pérdidas económicas en la producción de especies forestales (Pijut et al. 2011). Es por ello que el proceso de rizogénesis ha interesado a varias áreas de las ciencias y existen numerosas revisiones que muestran los avances en la biología de la formación de raíces adventicias (De Klerk 2002, Geiss et al. 2009, Kurepin et al. 2011, López-Villalobos et al. 2011, Oinam et al. 2011). Dentro de las investigaciones realizadas, los estudios histológicos tanto de los ejes como del desarrollo de los primordios radicales constituyen una base de conocimiento para entender la formación de las raíces adventicias (Gahan y George 2008, Oinam et al. 2011).
El corte transversal del entrenudo de regiones apicales de los brotes rejuvenecidos de plantas madres del minijardín clonal de P. alba mostró haces vasculares en diferentes estadios ontogénicos. Esto fue debido a variaciones en el patrón de diferenciación de xilema y floema y al inicio del crecimiento secundario que se produce muy próximo a las regiones apicales en la mayoría de las especies leñosas (Philipson et al. 1971).
En el cortex de miniestacas de P. alba se observó un anillo de esclerénquima con la típica estructura encontrada en la mayoría de las dicotiledóneas leñosas (Duarte y Wolf 2005, Ló y Duarte 2011). En los brotes rejuvenecidos de P. alba el denominado anillo de esclerénquima estuvo formado por dos regiones: los casquetes de esclerénquima y las regiones entre casquetes. Las discontinuidades en el anillo de esclerénquima se completaron con parénquima y pueden diferenciarse en células de gruesas paredes celulares. Las discontinuidades producidas por la presencia de células de parénquima no lignificadas o con menor grado de lignificación, se dieron en determinados estadios de diferenciación y fueron disminuyendo hasta desaparecer hacia la base de la rama. Lo anterior fue descripto para otras especies leñosas (Amissah et al. 2008).
En las miniestacas de P. alba que enraizaron se observó que los primordios radicales atravesaban el anillo de esclerénquimaa través de las regiones entre casquetes de esclerénquima, es decir, se observó enraizamiento en la región del anillo con menor grado de lignificación. En P. chilensis (Caro et al. 2002) y en Olea europea (Peixe et al. 2007) el enraizamiento logrado se relacionó con discontinuidades en el anillo de esclerénquima ubicado internamente al cortex radical. Para las mismas condiciones de enraizamiento, miniestacas de P. alba que presentaron mayor grado de lignificación en esas regiones no enraizaron; resultado también observado en otras especies leñosas (Bastos et al. 2005, Amissah et al. 2008, Elbasheer y Elkalifa 2010). En Paulownia fortunei var. mikado, por otra parte, se reporta una excelente producción de raíces en miniestacas gracias a la falta de una barrera anatómica esclerificada (Stuepp et al. 2015). Aunque, en otras especies como Camellia sinensis L. el anillo de esclerénquima no se comportó como una barrera mecánica en el momento de producirse el desarrollo de las raíces adventicias (Koyuncu y Balta 2004).
Edwards y Thomas (1980) han sugerido que la presencia de una capa continua de esclerénquima podría actuar como una barrera fisiológica a la iniciación de las raíces adventicias más que como una barrera mecánica a la emergencia radical. Por otra parte, Amissah et al. (2008) han considerado que las diferencias en el proceso de enraizamiento estaban más relacionadas con la facilidad de iniciación de raíces que con restricciones a su emergencia debidas al anillo de esclerénquima.
El enraizamiento puede verse dificultado por cambios en el grado de maduración de los ejes o por condiciones ambientales (Meng et al. 2009, Rasmussen y Hunt 2010, Oinam et al. 2011, Pijut et al. 2011). La formación de raíces adventicias es un proceso organogénico complejo que está afectado por la interacción de factores endógenos, tales como las hormonas vegetales y los carbohidratos, y por factores ambientales tales como la luz (Sorin et al. 2005, Druegge et al. 2008, Ahkami et al. 2009, Bennett y Scheres 2010, Kitomi et al. 2011, Verstraeten et al. 2013). En relación con ello se conoce que las condiciones ambientales y prácticas de manejo modifican los porcentajes de enraizamiento logrados (Rasmussen et al. 2009, Regonezi et al. 2010). Algunos de estos cambios pueden modificar el grado de lignificación de los ejes (Hatzilazarous et al. 2009). Cuando estos cambios modifican el grado de lignificación de las regiones entre casquetes de esclerénquima, modifican consecuentemente, el porcentaje de discontinuidades en el anillo de esclerénquima (Amissah y Bassuk 2007, Amissah et al. 2008). Las diferencias entre estacas y miniestacas de P. alba con relación a la lignificación del anillo estuvieron relacionadas a la etiolación, causada por la ausencia de luz. La generación de materiales juveniles a través de técnicas in vitro o usando métodos ex vitro como la etiolación (De Klerck 2002) condujeron a una exitosa formación de raíces adventicias. La etiolación se caracteriza por alteraciones fisiológicas asociadas a una reducción de la lignificación de los tejidos (Amissah y Bassuk 2007, Amissah et al. 2008). En P. alba la etiolación, al reducir la lignificación ha incrementado el porcentaje de discontinuidades del anillo de esclerénquima y las tasas de enraizamiento, tal como fuera planteado para otras especies leñosas (Amissah y Bassuk 2007, Amissah et al. 2008). Borges et al. (2011) han observado, en híbridos de Eucalyptus globulus, que las miniestacas obtenidas de la región apical de los brotes presentaban menor lignificación de los tejidos y tasas mayores de enraizamiento, como se observó en este trabajo.
Con relación a las concentraciones de IBA en el porcentaje de enraizamiento de las miniestacas, no se han observado diferencias significativas, posiblemente debido al grado de juvenilidad y lignificación de estos materiales, que permitieron altas tasas de enraizamiento, independientemente de la concentración de hormonas utilizadas. Lo mismo fue observado por Araujo Vieira de Souza et al. (2009) en el porcentaje de enraizamiento de cedro australiano, con la utilización de la técnica de miniestacas.
Con el aumento del IBA se registró un aumento de tasas de las variables analizadas hasta un punto óptimo y posterior a este punto el aumento de la dosis de IBA ocasionó una reducción de las tasas. Comportamiento similar fue observado en clones de Eucalyptus (Goulart et al. 2008). Concentraciones más elevadas de IBA también ejercieron un efecto negativo en el enraizamiento de estacas herbáceas de P. alba (Oberschelp y Marcó 2010).
CONCLUSIÓN
Al disminuir el grado de lignificación mediante rejuvenecimento (a través de la técnica de miniestacas) y etiolación de los propágulos utilizados para la propagación, mayores fueron las discontinuidades en el anillo de esclerénquima y mayor el grado de enraizamiento logrado en miniestacas de esta especie nativa. Lo mismo fue observado con miniestacas en relación a estacas. Esto señala la influencia del grado de lignificación en la propagación vegetativa de esta especie. Cuanto menor es el grado de lignificación mayor es la tasa de enraizamiento.
La utilización de la técnica de miniestacas, asociada a las técnicas de etiolación empleadas posibilitaron tasas de enraizamiento viables comercialmente, se muestran promisorias para la propagación vegetativa de P. alba y posiblemente de otras especies nativas de difícil enraizamiento.

