











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCTION
The genus Plebeia Schwarz comprises small social bees belonging to the tribe Meliponini (Apidae) or stingless bees. It is one of the most diverse Neotropical genera, with about 40 described species and many as of yet undescribed species (Rasmussen & Cameron, 2010). The genus is mostly characterized by small body size (3-6 mm), yellow markings on face and mesosoma, integument with sparse punctuation, mostly shiny and smooth (in contrast to e.g. Paratrigona Schwarz) and a depressed rim along the posterior margin of the inner surface of the hind tibia (Michener, 1990, 2007; Melo, 2016). Although morphologically quite homogenous, molecular studies suggest that the genus is polyphyletic with respect to Lestrimelitta Friese and Friesella Moure (Rasmussen & Cameron, 2010). Recently, Werneck (2016) in a comprehensive work analysing Plebeia, reaffirmed the polyphyly of the large genus Plebeia and the monophyly of the smaller genera Lestrimelitta and Friesella.
Species of Plebeia are generally very abundant and occur in most of the Neotropical region, from Nuevo León, Mexico to Buenos Aires and San Luis, Argentina, representing also the distributional boundary of the stingless bee tribe in the Neotropical región (Roig-Alsina & Alvarez, 2017).
In this work we describe and figure a new species of Plebeia from the Argentine Yungas and provide details of its nesting behavior. The Yungas extends along the eastern slopes of the Andes, as a narrow strip, from Venezuela to the northwest of Argentina, in the provinces of Jujuy, Salta, Tucumán and Catamarca. Altitudinally, it extends between 500 and 2,500 to 3,500 m (Cabrera & Willink, 1980), with the argentine stingless bee fauna of this region being the least known of the country (Roig-Alsina et al., 2013).
MATERIAL AND METHODS
Morphological terminology follows that of Michener (2007), except for surface sculpture, which follows Harris (1979), and the descriptive format generally follows that of Melo (2016). Abbreviations used in the descriptions are F, S, T and pd for flagellomere, metasomal sterna, metasomal terga and puncture diameter, respectively. External morphological features were studied using a Nikon SMZ 745T stereomicroscope. Photographs were taken with a Canon T3i digital camera attached to a stereomicroscope. Digital images were afterward stacked using the CombineZM open source software. The distribution map was produced with the program SimpleMappr (Shorthouse, 2010). The measures in the descriptions are for the holotype, with ranges of standard deviation from five of the paratype specimens provided in parentheses. Material examined for this study is deposited in the following collections: CRCD, Claus Rasmussen Collection, Aarhus University, Denmark; IBSI, Instituto de Biología Subtropical, Puerto Iguazú, Argentina; IFML, Fundación Miguel Lillo, San Miguel de Tucumán, Argentina; MACN, Museo Argentino de Ciencias Naturales "Bernardino Rivadavia”, Buenos Aires, Argentina; MLP, Museo de La Plata, La Plata, Argentina; NHMUK, The Natural History Museum, London, UK and SEMC, Natural History Museum University of Kansas, Kansas, USA.
This article has been registered in the Official Register of Zoological Nomenclatura (ZooBank) as LSlDurn: lsid:zoobank.org:pub:6B7E13B9-9C73-413E-938D-D8A FF4975438
RESULTS
Plebeia mansita Alvarez & Rasmussen sp. nov.
(Figs. 1-3 and 5-9)
Plebeia intermedia (Wille): Flores et al., 2015: 515-529 (misidentification)
Plectoplebeia nigrifacies (Friese): Alvarez et al., 2016: 72 (misidentification)
Plebeia sp.: Flores et al., 2018
DiagnosisPlebeia mansita sp. nov. is distinguished from other species of the genus Plebeia by the following combination of characters: relatively large body size (~4.9 mm), supraclypeal area and scape dark brown to black, malar space wide and yellow wide band on posterior half of scutellum. In South America only two described species are of similar large size: P remota (Holmberg, 1903) and P saiqui (Friese, 1900). Plebeia remota can be easily distinguished by the narrow malar area, coloration of the supraclypeal area and long yellow paraocular marks. The long malar space is shared with P saiqui, from which it is separated by the presence in P saiqui of dull yellow spots on the body, black scutellum or at most with small dull yellow spots on the posterior margin, and the shape of preapical teeth (Fig. 4) (first preapical tooth widest and second preapical tooth more acutely pointed than in P mansita sp. nov.).
DescriptionHolotype worker. Approximate body length 4.9 (± 0.22) mm; maximum head width 2.0 (± 0.06) mm; forewing length 5.1 (± 0.12) mm; maximum width of mesoscutum 1.35 (± 0.05) mm. Color: Integument predominantly dark brown to black, except following yellow areas: narrow strip on the lower paraocular area (Fig. 1); small triangular spot on lower lateral angle of clypeus; interrupted band on pronotum and small spot on pronotal lobe; strip along each side of mesoscutum; dorsal portion of axilla; wide band on the posterior half of scutellum (Fig. 2); and small spot on apex of fore femur and base of fore tibia. Labrum yellowish, with basal narrow dark band; mandible reddish brown, except for black base and apex brown (Fig. 3); central portion of metanotum, mediotarsi, distitarsi and inner surface of hind basitarsus light brown. Wing veins and pterostigma brown; wing membrane light brown (Fig. 5). Pubescence. Predominantly yellowish to light brown; tibia and tarsi with dark brown setae. Clypeus densely covered with short, whitish and plumose pubescence, sparser intermingled erect long and light brown setae about 0.08 mm in length. Frons with white, short and dense plumose pubescence, intermingled with sparser simple and longer light brown setae, longest setae on upper frons and vertex 0.14-0.20 mm in length; erect setae on scape short, slightly shorter than diameter of scape (0.08 : 0.12); gena with a dense pile of short decumbent plumose pilosity, partially obscuring integument. Scutum covered with a dense pile of short simple pubescence and sparser intermingled longer hairs; hairs along anterior margin mostly plumose, longest about 0.20-0.28 mm, those on remainder of sclerite simple and shorter (0.06-0.12 mm); longest setae on scutellum 0.20-0.24 mm in length. Lateral portion of mesepisternum with short, pale and plumose pubescence intermingled with sparser simple and longer light brown hairs, longest hairs on ventral region about 0.16-0.20 mm in length, metepisternum and lateral portion of propodeum with similar vestiture, shorter plumose pilosity denser compared to mesepisternum. Propodeal triangle glabrous. Tergum 1 glabrous, T2 with only sparse minute hairs along hind margin, pubescence denser and longer on lateral portion; T3-T5 with pubescence restricted to posterior margin, forming complete bands of relatively short, mostly erect, simple setae, longest setae 0.06-0.12 mm; pubescence of T6 similar to T3-T5, but more abundant, completely covered, lacking plumose hairs. Sternawith erect simple hairs, S1 and S2 with a curved apex; longest setae on S2 0.2-0.3 mm in length, S6 with abundant plumose hairs on central portion. Integumental surface: Mostly smooth and shiny, punctuation relatively thin and abundant on the head and mesosoma. Clypeus and supraclypeal area rugulose and somewhat dull, finely punctured, punctures separated by 1-2 pd. Frons with coarse punctures; separated by 2-3 pd, smooth and shiny integument between the punctures on mid portion; punctation on lower paraocular area and portion near orbits less dense (3-4 pd); ocello-ocular area and vertex finely rugulose and somewhat dull. Punctures on gena similar to those on frons, but denser (about 0.5 pd). Punctures on mesoscutum coarser than those on frons, separated by 1-2 pd, interspaces mostly smooth and shiny, except fine reticulation along anterior margin; scutellum with punctures slightly sparser than those of mesoscutum, interspace smooth and shiny, except finely rugulose integument on posterior margin. Disc of mesepisternum shiny and smooth, punctures separated by 2-4 pd, superior and posterior margins with punctation denser and integument rugulose-reticulate; metepisternum rugulose along anterior margin. Lateral portion of propodeum shiny but distinctly reticulate, puctures separated by 0.5-1 pd; surface of posterior portion finely reticulate. Metapostnotum striate along anterior margin, remainder finely reticulate to imbricate. Terga mostly shiny, with fine reticulation on apical margin of T1, apical half of T2 and visible portions of T3-T6. Structure: head about 1.2x (2 ± 0.06 : 1.64 ± 0.04 mm) wider than long and 1.3x (2 ± 0.06 : 1.5 ± 0.03 mm) wider than clypeo-ocellar distance; inner orbits converging below (upper to lower interorbital distance, 1.33 ± 0.03 : 1.07 ± 0.04 mm); maximum inter-obital distance about 1.14x greater than length of eye (1.45 ± 0.04 : 1.27 ± 0.03 mm); frontal line distinct, sulciform; clypeus 1.95x wider than long (0.48 ± 0.02 : 0.94 ± 0.01 mm); F2 wider than long (0.16 : 0.13 mm); F3 as long as wide (0.14 : 0.14 mm); minimum distance between clypeus and eye about 0.5 times the width of F2 (0.07 : 0.13 mm); malar space about 0.5 times as long as F3 width (0.08 ± 0.008 : 0.15 ± 0.01 mm); eye about 2.8 longer than its maximum width (1.27 ± 0.03 : 0.45 ± 0.03 mm), in lateral view as wide as gena (0.2 ± 0.03 : 0.18 ± 0.04 mm). Inter-tegular distance slightly greater than mesoscutum length (0.7 ± 0.04 : 0.64 ± 0.04 mm); scutellum about 2.1x wider than long (0.88 ± 0.03 : 0.42 ± 0.02 mm). Hind tibia 2.9x longer than wide (1.78 ± 0.03 : 0.62 ± 0.02), hind basitarsus 1.8x longer than wide (0.92 ± 0.02 : 0.46 ± 0.03). Forewings longer than body length, marginal cell about 4x longer than wide, hind wings with six hamuli. Mandible with two preapical teeth separated by a narrowly concave emargination, first preapical tooth rounded, second pointed (Fig. 3). Propodeal spiracle elongate, about 3.7 times longer than wide.

Figs. 1-5: Worker of Plebeia mansita sp. nov. 1, head in frontal view; 2, mesosoma in dorsal view; 3, detail of mandible; 4, detail of mandible of Plebeia saiqui; 5, lateral view. Scale bars 1 and 2 = 0.5 mm; 3 and 4 = 0.25 mm; 5 = 1 mm.
Male. Unknown.
Queen. Unknown.
Variation: Yellow spots on the clypeus and paraocular area quite variable, generally present, in some specimens poorly developed or absent. The number of hamuli variable, specimens examined have from five to seven.
Type Material: Holotype; worker, MLP, N° 5735/1: ARGENTINA, Salta, Baritú, 22-IX-2011, (22° 28’54’’S; 64°45’39”W, 1546 m), F. Flores. Paratypes; 2 workers, with same data as holotype (CRCD); 2 workers, with same data as holotype (IBSI); 1 worker, with same data as holotype (IFML); 2 workers, with same data as holotype (NHMUK); 2 workers, with same data as holotype (SEMC); 18 workers, with same data as holotype (MLP); 12 workers, Jujuy, San Lucas, Depto. Valle Grande, 16-XI-2010, 16- XII-2010, (-23.533S; -65.083W, 1896 m), F Flores (MLP); 12 workers, Jujuy, Potrero de Yala, (-24.1210; -65.4644), 20-IV-2015, R. González V. (MACN); 1 worker, Salta, Oran, R. Nac. 57 Km. 21, El Chorro, 1130 m., 28-X-1978, P. Fidalgo (IFML); 2 workers, Salta, San Andrés, Depto. Orán, 15-I-2007, (-23.100481; -64.871035, 1814.8 m), F Zamudio (MLP).
Etymology: The epithet refers to the common name used by local people in Baritú, El Lipeo and Los Toldos, “mansita", due to the docile behavior of the bees when disturbed or handled.
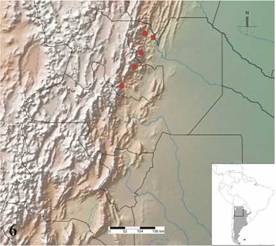
Distribution: Argentina, provinces of Jujuy and Salta, restricted to the biogeographic province of Yungas at altitudes above 1,000 m (Fig. 6).
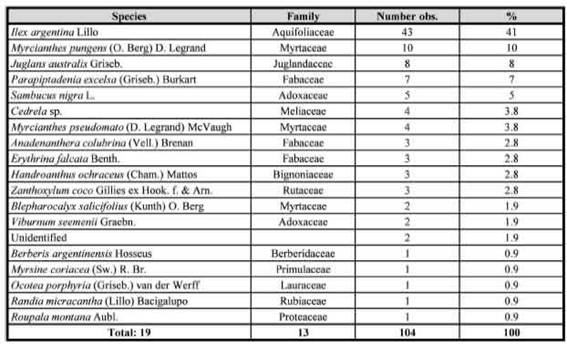
Nesting biology: Nests of Plebeia mansita sp. nov. were located inside pre-existing cavities of live tree trunks. One of us (F.F.F.) studied the nesting substrates of 104 colonies in different locations in the provinces of Jujuy and Salta. Plebeia mansita sp nov. occurred in 19 tree species of 13 families, with Ilex argentina Lillo (Aquifoliaceae) being the most used (41%), followed by Myrcianthes pungens (O. Berg) D. Legrand (Myrtaceae) (10%), Juglans australis Griseb. (Juglandaceae) (8%), Parapiptadenia excelsa (Griseb.) Burkart (Fabaceae) (7%), Sambucus nigra L. (Adoxaceae) (5%), Cedrela sp. (Meliaceae) and Myrcianthes pseudomato (D. Legrand) McVaugh (Myrtaceae) (3.8% each) (see Table I). Their distinct nest entrances were located at any height between 5 cm and 6 m above ground. The nest entrance is, contrary to most other species of Plebeia, not formed by a small or narrow cerumen tube. The entrance is instead wide and highly variable as it traces the shape of the host tree crevice. Most of the surface of the crevice can be coated and closed by dark propolis and cerumen, forming one to three smaller holes leading into the brood area (Fig. 7). In these orifices, numerous bees can enter at the same time. In general, 10 to 25 guard bees are also present at the nest entrance (Figs. 7and 8). The lower part of the external nest entrance usually presents a characteristic dark stain (Fig. 8, arrow) of variable extension that is apparently produced by the bees. This structure could function as a visual or chemical signal for landing. The brood chamber was covered by a thick involucrum, consisting of 5 to 10 layers of cerumen. The brood cells were arranged in combs separated by pillars (Fig. 10). Storage pots were arranged in groups of about 75 (n = 3 nests), supported by the surrounding involucrum. The pollen and honey pots measured 15-23 mm tall (mean = 19) by 9-13 mm in maximum width (mean = 11), both had similar ellipsoid shape. Workers of P. mansita sp. nov. are very docile, with the bees remaining active during close inspection. According to the local people surveyed, the nests of P. mansita sp. nov. produce 0.25 to 8.00 liters (mean = 2.40) of honey (n = 56 nests). It is a significant production of honey and considering the docile behavior of the species, it makes it a good candidate for rational breeding.

Figs. 7-10: Nesting biology of Plebeia mansita sp. nov. 7 and 8, nest entrances; 7, on a tree trunk of Ilex argentina Lillo (Aquifoliaceae); 8, on a tree trunk of Juglans australis Griseb. (Juglandaceae); 9, nest entrance showing the dark stain (red arrow); 10, brood chamber, showing the layers of cerumen of involucrum and the combs of brood cells separated by pillars.
DISCUSSION
Plebeia mansita sp. nov. shares numerous characteristics with the distinct genus Plectoplebeia with which it has been confused (Alvarez et al., 2016). Superficially it resembles Plectoplebeia by presenting a similar coloration, pubescence and integument sculpture. They also agree in the relatively large body size, wide distance between the clypeus and the eye, an elongate hind tibia, high number of hamuli and relatively long wings. However, P. mansita sp. nov. differs mainly in the shape of the scutellum, the elongate propodeum and metaposnotum and the rectangular T1. It is now clear that those specimens are distinct and represent the new species of Plebeia described herein. Although there is no morphological evidence to differentiate the two independent clades of Plebeia (Rasmussen & Cameron, 2010), P mansita sp. nov. would appear to be related to the larger clade that has Lestrimelitta and Friesella as sister groups. Under the erroneous name Plebeia intermedia, Flores et al. (2015) carried out a botanical characterization of P mansita honey, identifying 54 pollen types, especially of native plants of arboreal habit. Flores et al. (2018) provided comprehensive information on the uses of this species by local people and features of nesting habits.
In the key to Argentine Plebeia species by Alvarez et al. (2016), P. mansita sp. nov. partially runs to the first couplet, which is here amended as follows:
1. Relatively large bees; length of the anterior wing 4.5 to 5.2 mm; head width ~ 2.0 mm; vertex and mesoscutum with brown pubescence..........................2*
-. Bees of smaller size, length of the anterior wing 2.8 to 3.6 mm; head width from 1.4 to 1.8 mm; vertex and mesoscutum with whitish pubescence..................2
2*. Supraclypeal area black; malar space wide, about 0.5 times width of F3.......................P. mansita sp. nov.
-. Supraclypeal area yellow, pale, or brownish; malar space narrow, about 0.3 times width of F3.................................................P. remota (Holmberg)
ACKNOWLEDGMENTS
We are grateful to anonymous reviewers for insightful suggestions and comments on the manuscript. F.F.F. wants to thank the residents of El Lipeo, Baritú, Ocloyas and Valle Grande, in particular Nieves Grimaldo, Josefa Vilte, Concepción and Bonifacio Aramayo, Onorato and Mirta Cardozo, Carlos Aparicio, Eliseo Ontiveros, Miriam Méndez and their families, who kindly assisted us and shared their experience and wisdom as well. L.J.A. would like to thank Ana Maria Marino de Remes Lenicov and Barbara Defea for their help in obtaining the photographs. This work was partially supported by PICT 2016-1846. Financial support to L.J.A. and F.F.F. was provided by the Consejo Nacional de Investigaciones Científicas y Técnicas, Argentina (CONICET).

