










Serviços Personalizados
Journal

Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkRevista Argentina de Ciencias del Comportamiento
versão On-line ISSN 1852-4206
Rev Arg Cs Comp. vol.8 no.1 Córdoba abr. 2016
ARTICULO ORIGINAL
Analysis of post-agonism reconciliation in a captive group of the wedge-capped capuchin monkey, Cebus olivaceus: A pilot study.
Tárano, Zaidaa & Flores, Ruth M.ab
a Laboratorio de Comportamiento Animal, Instituto de Biología Experimental, Facultad de Ciencias, Universidad Central de Venezuela, Venezuela.
b Foothill College, Los Altos Hills, California, USA.
Enviar correspondencia a: Tárano, Z. E-mail: zaida.tarano@ciens.ucv.ve
Abstract
Agonism was often associated to an increase in inter-individual distances in social individuals; however, many evidences indicate that individuals affiliate after a dispute ( “reconciliate”). The post-agonism behavior of individuals in a captive troop of the wedge-capped capuchin monkey was described and conciliatory tendency (CT) was estimated by using the PC-MC (Post-Conflict – Matched Control) method; here PA-MC (Post-Agonism – Matched-Control). Males initiated more agonistic interactions toward females than to other males while females avoided initiating them toward males. Interestingly, females were more prone to initiate reconciliation than males, both toward males (CT: 0.20-0.42) and toward other females (CT: 0.24-0.25). Overall, the CT in the group ranged from 0.127 to 0.198. We compared our results with those from other capuchins and propose that reconciliation may not be fundamental in maintaining social cohesion in Cebidae; nonetheless, due to our small sample, this is a preliminary conclusion.
Key Words: Reconciliation; Capuchin Monkey; Agonism; Conciliatory Tendency
Resumen. Análisis de la reconciliación post-agonismo en un grupo en cautiverio del mono capuchino común, Cebus olivaceus: Un estudio piloto. El agonismo frecuentemente se asoció con el incremento distanciamiento interindividual en grupos sociales, pero las evidencias indican que los individuos se afilian después de una disputa (“se reconcilian”). Las conductas post-agonistas se describieron en un grupo de monos capuchinos en cautiverio y se calculó la tendencia conciliatoria (TC) utilizando el método pareado PC-CP (Post-Conflicto – Control Pareado); aquí PA-CP (Post-Agonismo – Control Pareado). Los machos iniciaron más interacciones agonistas hacia las hembras que hacia otros machos, mientras que las hembras evitaron dirigirlas a los machos. Interesantemente, las hembras iniciaron más reconciliaciones que los machos tanto hacia machos (TC: 0,20-0,42) como hacia hembras (TC: 0.24-0.25). Globalmente, la TC del grupo varió entre 0,127 y 0,198. Comparamos nuestros resultados con los de otros capuchinos y proponemos que la reconciliación podría no ser fundamental en el mantenimiento de la cohesión social en Cebidae; no obstante, debido al tamaño de nuestra muestra, esta conclusión es preliminar.
Palabras claves: Reconciliación; Mono Capuchino; Agonismo; Tendencia conciliatoria
Recibido el 8 de Octubre de 2015; Recibida la revisión el 5 de Noviembre de 2015; Aceptado el 17 de noviembre de 2015.
Editaron este artículo: Mariana Bentosela, Soledad Sartori, Eliana Zamora y Estefanía Caicedo.
1. Introduction
Group living has evolved in many taxa from invertebrates to mammals because it confers a number of advantages such as improved food acquisition, resource defense, anti-predatory defense and communal care of offspring among others; nevertheless, group living also entails significant costs (Alexander, 1974). With a few exceptions sociality is the norm among primates (Smuts, Cheney, Seyfarth, & Wrangham, 1986). In primate´s groups, social relationships are fundamental in maintaining group cohesion (van Hooff & van Schaik, 1992; Kappeler, 1993; Welker, Becker, Höhmann, & Schäfer-Witt, 1987), and are behaviorally evidenced through affiliative and agonistic behaviors (Welker et al., 1987). Affiliative behaviors are pacific acts between two or more individuals such as sit beside, sniff, lick, touch, allogroom, embrace and kiss. Agonism refers to behaviors expressed in situations of competition and usually involve intimidation, fight and submission, through ritualized aggressive displays, groans, shouts, hair erection, chases, pushes, hair pulling, bites and escalated fight; therefore conflict of interest among individuals is inferred through agonistic behaviors (Welker et al., 1987).
Most of the outcomes of agonistic interactions involve an increase in the inter individual distance, but after de Waal & van Roosmalen´s (1979) observations on a captive colony of chimpanzees, affiliation after a dispute between former opponents has been recorded often in several species from birds to primates (references in, Cools, van Hout, & Nelissen, 2008; Logan, Emery, & Clayton, 2013; Polizzi di Sorrentino, Schino, Visalberghi, & Aureli, 2010; Silk, 2002). This phenomenon has been called reconciliation, and it does not imply any emotional attitude or the resolution of the underlying unknown conflict between the individuals. Reconciliation has been operationally defined as friendly (affiliative) reunions between former opponents occurring soon after an aggressive interaction (de Waal, 2000). Several non-mutually exclusive hypotheses have been postulated to explain short and long term consequences of reconciliation (reviews in, Arnold & Aureli, 2007; Silk, 2002). The stress reduction hypothesis (reconciliation reduces physiological stress) and the pacific intention or uncertainty reduction hypothesis (reconciliation reduces the uncertainty about the nature of future interactions) are the most robust among short term-benefits hypothesis; while the relationship quality or valuable relationships hypothesis (reconciliation preserves valuable relationships) is the most robust among several long term-benefits hypothesis (reviews in, Arnold & Aureli, 2007; Silk, 2002).
Post-agonism affiliation has been mostly studied in old world primates and great apes, with relatively scarce research in Neotropical primates. Available information has revealed striking differences among them in the type of conciliatory behaviors (e.g., approach without contact, extensive physical contact, embraces, kisses, anogenital inspection, allogrooming) and in the time elapsed between the end of the agonistic interaction and the pacific interaction (Arnold & Aureli, 2007). Conciliatory tendencies (CT) also vary strongly from great apes (de Waal, 2000) to small arboreal primates (e.g., Daniel, Santos, & Cruz, 2009; Kappeler, 1993; Leca, Fornasieri, & Petit, 2002; Manson, Perry, & Stahl, 2005; Palagi, Antonacci, & Norscia, 2008; Verbeek & de Waal, 1997; Visalberghi & Fragaszy, 1995; see also Thierry et al., 2008 for results on semiterrestrial species of Macaca). This great variability justifies continuing research in this topic, and specifically in the new world family Cebidae. Some research has been carried out on post-agonism behaviour in two species of Cebidae, Sapajus apella (Daniel et al., 2009; Perry, 1995) and C. capucinus (Leca et al., 2002; Manson et al., 2005; Verbeek & de Waal, 1997; Visalberghi & Fragaszy, 1995) indicating relatively low reconciliation tendencies.
This is a pilot study in which we analyzed post-agonism behaviors in a troop of captive wedge-capped capuchin monkeys, Cebus olivaceus that has been extensively observed by López and Tárano (2009) and Tárano and López (2015). In the previous studies, these authors described the behavior of one semicaptive and two captive groups and found that agonist behaviors were less frequent while affliliative ones were as frequent in the study groups as in the wild (Robinson, 1981), but shorter and less intimate (less body contact). López and Tárano (2008) reasoned that reduced agonism together with relatively high affiliation would be tension-management mechanisms (Nieuwenhuijsen & de Waal, 1982) resulting from the confinement of these highly mobile animals. In the present study, we focused our observations in one of the previously studied groups, specifically in the captive group with the lowest density and the least distorted group composition (age-sex composition), which was also the easiest to observe.
2. Method
2.1. Study Subjects and Site
The wedge-capped capuchin monkey Cebus olivaceus Schomburgk, 1848 (Cebidae, Cebinae) is a medium-sized arboreal monkey of approximately 2.5-2.8 kg, found in South America from Venezuela to the southern Amazon Basin in Brazil (Eisenberg & Redford, 1989). Wild groups are formed by 8-36 individuals depending on the habitat, 17% adult males, 33% adult females, 41% young and 9% infants (Fragaszy, Visalberghi, & Fedigan, 2004). Females are philopatric while males switch between groups throughout their lives. Each group has an alpha male and an alpha female but the hierarchy is not clear below the top individuals (Fragaszy et al., 2004; Robinson, 1988). In the wild, most agonistic interactions occur with or among the high rank individuals, typically over food, and the low rank males avoid interacting with the higher rank ones (Fragaszy et al., 2004).
We conducted the study at the recreational zoological park Generalísimo Francisco de Miranda (10º25’50”N, 66º58’03”W) in Caracas (Venezuela). The monkeys are restricted to an outdoor enclosure limited by a shallow lagoon which included two concave islands (245 m2 approx.) connected by a wooden bridge. The larger island has a subterranean cage in which the animals rest during the night. The islands have several trees, a cement feeder on the floor, several wooden cages, ropes, rope bridges and several poles. The diet consists of fruits (banana, papaya, cantaloupe, fresh corn); boiled eggs and cooked-chicken wings are also provided once a week.
At the moment of the study, the group had 17 individuals: 5 males (3 adults, 2 sub-adults), 5 females (3 adults, 2 sub-adults), 5 juveniles and 2 neonates born during the study period (juveniles and neonates were not included in the observations). The proportion of adult males to females was 1:1 and was biased toward males with regards to that observed in the wild (typically 1 adult male: 2 adult females) (Fragaszy et al., 2004; Robinson, 1988). The individuals were identified by natural markings such as colour patterns, scars and differences in size that were easily observed from a distance. The composition of the group had remained stable during five years prior to the present study (M. CH. López. pers. comm.) with the exception of several new births and introduction of an adult male after the death of the dominant one three years earlier; this new male soon reached the dominant position and remained so during the study period (A. Quintero, pers. comm.). The group had a dominant pair, several subdominants of similar rank and few lower rank individuals (R. Flores, unpublished data). Within-sex hierarchies indicated that, among males, there were two subdominants of similar rank, followed by a third rank individual and a bottom one (Figure 1A). Among females, there was a subdominant (probably the daughter of the dominant female), a third rank female and two bottom rank females (Figure 1B), one of which was the oldest female and the least active individual in the group; the other bottom rank female was probably her daughter. The dominant individuals receive the highest proportion of affiliative interactions, and mainly from females, which in addition, are more prone to initiate affiliative interactions than males, and mainly toward other females (López & Tárano, 2008). Mid-rank males give a disproportionate amount of affiliative interactions toward the dominant male (R. Flores, unpublished data). These patterns of dominance and affiliation are qualitatively identical to those described in wild groups (O´Brien, 1991).
The study was conducted with authorization of the zoo´s managing board (Ing. Juan Rodríguez, Ing. Ramón Alis Rojas, & Arq. Norma Marín) and required no other permission since individuals were nor disturbed in any way, but merely observed from a distance much as visitors do.
Figure 1.
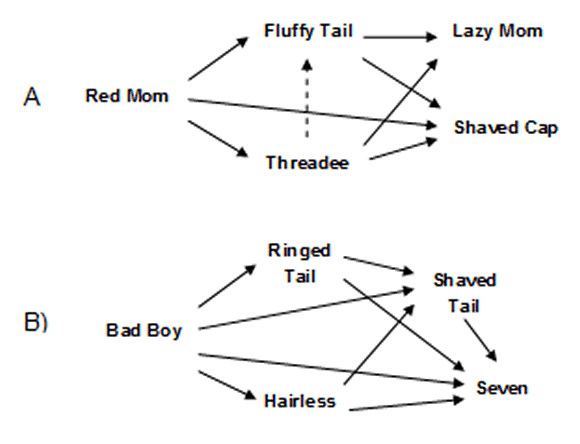
Dominance relataionships based on interacctions lost and won by each individual (R.M. Flores, unpublished data). A) Females. B) Males. The relative rank of Threadee and Fluffy Tail (in dots) is dubious because it is based in one interaction.
2.2. Observation Methods
We performed the observation during 6 months spanning from April until September 2010, during three days a week, from 09:00 to 12:00 hrs and from 14:00 to 16:30 hrs, for a total of 70 effective days and 64 hours of video analysis (see below). This sampling effort was similar to that reported by Robinson (1981) for a wild troop. To estimate the likelihood of reconciliation we used a combination of all occurrences sampling and focal sampling (Altman, 1974). During all occurrences sampling, we recorded the behaviors of all individuals in the group, either by remaining still at one location (provided all the individuals were visible from that position) or by slowly walking around the pit watching all the individuals at once. When an agonistic interaction was initiated, we focused a video camera (Sony Handycam CCD-TR517 8mm, supported by a tripod) toward the interacting individuals, recorded the initiator (aggressor) and the receiver (victim) of the interaction, the type of agonistic behavior and its duration. We operationally defined the onset of an aggressive interaction as an approach toward another individual involving a hasty change in activity, movement direction or speed, accompanied by subtle modifications in posture, facial expression or vocalizations indicative of aggressive intention. An interaction could involve only one behavioral event or several subsequent events of the same type or not. We defined the interaction ending as a separation of the individuals larger than 10 arms´ lengths or a cessation of the agonistic behaviors for a period longer than 30 seconds. Then, we began a focal observation on one of the former opponents during 10 minutes. During this period (post-agonism, PA; post-conflict in general literature on this topic), we observed any affiliative interaction between the individuals, and when it occurred, we measured the time elapsed from the end of the agonistic interaction until the beginning of the first affiliative one, this is called the response time (Silk, 2002; de Waal, 2000); additional interactions were also recorded. We preferred the term agonism instead of conflict because the latter refers to the underlying, typically unknown, source of the competitive differences between individuals while the former simply refers to the type of interaction. To determine whether there was reconciliation we performed another 10-min observation on either of the former opponents, approximately at the same hour of the PA observation and under the most similar conditions but provided that there has been no agonism between them at least 2 hours prior the observation (matched control observation or MC). During MC, we also measured the time elapsed from the beginning of the observation until the occurrence of the first affiliative interaction and additional interactions. This method, known as PC-MC (PC means post-confrontation, here referred as post-agonism), was introduced by de Waal and Yoshihara (1983).
With these data, we estimated the likelihood of reconciliation through: the comparison of the distribution of the response times in the PA and MC periods, and the calculation of a conciliatory tendency (CT). The response time estimates the latency to reconciliation between the former opponents, therefore the coincidence of the PA-MC distributions would indicate a random pattern and that reconciliation does not occur. On the contrary, when the distribution of the response times during PA is biased toward lower values than that during MC, reconciliation is said to occur (Silk, 2002). To calculate CT, we compared the response times within each PA-MC paired observation, and classified each pair as neutral (the response time was equal in both observation periods), attracted (the response time was shorter in PA than in MC) and dispersed (the response time was longer in PA than in MC). Then, we calculated the CT as: (Attracted pairs – Dispersed pairs) /N, with N being the total number of PA-MC pairs (Veenema, Das, & Aureli, 1994). The CT ranges from -1 to 1 and it is analyzed qualitatively.
2.3. Statistical Analysis
First, we analyzed whether there was association between the sex of the individuals and the probability of initiating an agonistic interaction or an affiliative interaction post-agonism with a c2, grouping categories if necessary to fulfill the requirement of having ≥ 5 observations in at least 80% of the cells (Sokal & Rohlf, 1995 ). We also analyzed the association between the role (receiver or aggressor) and the probability of initiating the affiliative interaction post-agonism. When significant results were found, we performed a standardized residuals analysis to determine which cells were responsible for the results, this is, to establish which cells significantly deviate from random expectation, with a Zcrit of ± 2.57 (α = .01).
To test for differences in affiliation during PA and MC we compared the duration and frequency of the affiliative interactions with a Wilcoxon matched pairs test. We also compared the distributions of the response times with a c2 goodness-of-fit test. In addition, we compared the number of attracted and dispersed pairs per dyad with the paired Wilcoxon sign test. All the analysis were performed with Statistica 10.0 and Microsoft Excel 2007.
3.1 Agonistic Interactions
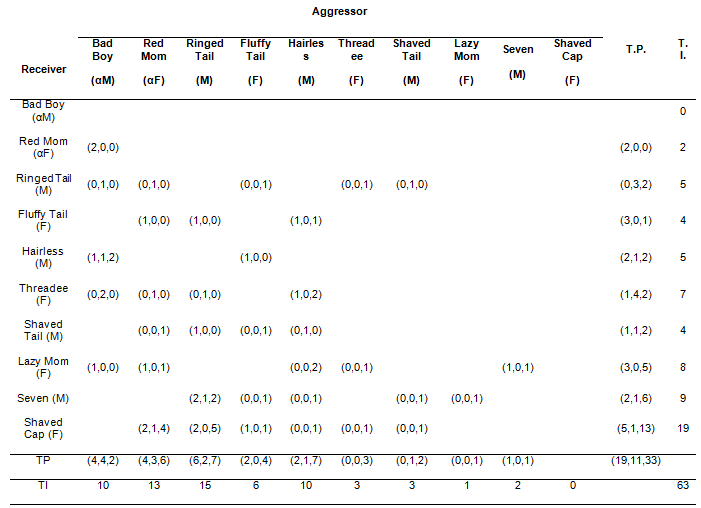
We registered 63 agonistic interactions during 64 hours of video analysis, corresponding to 85 agonistic events (several interactions consisted on more than one event) most of which were positively indentified (n = 77). We identified four types of agonistic behaviors (events): chase (n = 25), threatening approach (n = 24), physical attack (n = 20) and steal food (n = 8). Chases were runs on-all fours or jumping (hind legs), involving climbing onto branches during the chase, behind another individual with or without vocalizations. In threatening approaches, one individual moved toward another with dorsum hair raised, tail raised; mouth opened showing teeth, with or without vocalizations. Physical attacks involved physical contact such as hair pulling, pushing and pulling legs or tail and biting, usually with loud vocalizations uttered by both opponents. Steal food involved one individual slowly approaching another, rapidly taking away an item of food and then running away; the receiver usually responded by chasing the other individual. With regards to the sex of the initiator (aggressor) and the receiver, most agonistic interactions were initiated by males toward females (male-female) (Table 1, only the first agonistic event included in the analysis, c2 = 9.32, p = .025, d.f. = 3; significant residual = +5.43, p < .01); male-male and female-female interactions were equally common and did not depart from random expectation; the least common (and less than expected) agonistic interactions were those initiated by females toward males (significant residual = -3.81, p < .01).
Table 1. First agonistic event recorded during agonistic interactions in the study group.
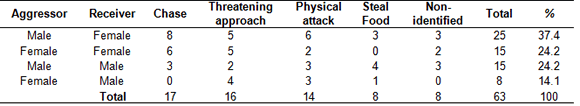
Significantly, males were prone to initiate interactions toward females (c2 = 9.32, p = .025, d.f. = 3; significant residual =+5.43, p < .01), while females avoided initiating interactions toward males (c2, significant residual = -3.81, p < .01)
3.2. Affiliative behaviors during PA-MC
We noted affiliative behaviors in 20 out of 63 PA observations and in 13 MC observations (Table 2). The first affiliative interaction between the former opponents was classified as: approach within arms´ length without contact (nPA = 11, nMC = 8), allogrooming (nPA = 5, nMC = 1), social play (nPA = 3, nMC = 4) and approach with contact (nPA = 1, nMC = 0). In approach without contact, one individual walked slowly on all fours toward another and sat besides him/her, laid on a side or on its back, or remained on all fours within arm´s length. Approach with contact was similar to the former but the approaching individual gently touched the other´s back, arm or shoulder with the dorsum or palm of its hand. In allogrooming, an individual poked the other´s fur or ears with fingers or teeth, scratched the skin or licked it. Social play involved mutual chases, tail pulling, jumps on each other´s back, jumps on hind legs facing each other, climbing or hanging on ropes while chasing another individual. We did not observe approaches with contact during control (MC) observations. Due to the small sample size which resulted in a large number of cells with zero observations in the contingency table, we did not perform statistical analysis but qualitatively, allogrooming seemed more common during PA than during MC (Table 2), while approaches without contact were similar in both periods.
Table 2. First affiliative interaction during PA (post-agonism) and MC (matched-control) sessions.
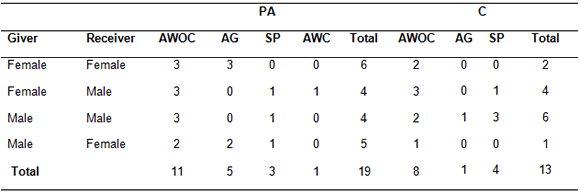
Note: AWOC: Approach without contact, AG: allogrooming, SP: social play, AWC: approach with contact.
Qualitatively, with regards to the sex, males and females initiated approaches toward either sex with similar frequency during PA (6 initiated by females and 5 by males), and were equally prone to initiate allogrooming (3 initiated by females and 2 by males), but both sexes directed these interactions toward females (no male received grooming). Approach with contact was observed only once and was initiated by a female toward a male. During MC sessions, approaches without contact were initiated mainly by females and received mainly by males, but female-female and male-male approaches were also observed. Male-male play was more common than female-male play while male-female and female-female play was not observed. Given the small sample size, we consider these qualitative associations with caution.
In addition, we observed secondary (additional) affliliative interactions in 19 PA periods and in 11 MC periods (usually one but occasionally up to six additional affiliations). In 12 PA periods, the second interaction was a repetition of the first and was initiated by the same individual (8 approaches without contact, 3 with contact and 1 social play). Only in one PA period was the second interaction identical to the first but initiated by the former receiver and in 2 cases, the second interaction was different from the first and initiated by the former receiver. In 7 MC periods, the second interaction was a repetition of the first and was initiated by the same individual (3 social plays, 3 approaches without contact and 1 allogrooming); in the other 4 periods, the interaction changed but was initiated by the same individual (approaches without contact). Altogether, the results indicate that former givers are more prone to initiate a second affiliative interaction than former receivers.
Affiliative interactions during PA and MC did not differ in duration (Wilcoxon: z = 0.42, p = .69), lasting on average PA = 0.47 ± SD 0.85 s (max = 4.03, min = 0.02) and MC = 0.53 ± SD 1.22 s (max = 4.65, min = 0.05). In addition, the frequency of affiliations during PA and MC observations was not significantly different (Wilcoxon matched pairs tests: z = 1.08, p = .29; PA = 0.63 ± SD 1.31 interactions per observation period, max = 7; MC = 0.40 ± SD 0.89 interaction per observation period, max = 4).
3.3. Response time and conciliatory tendency
Overall, 60% of the interactions occurred within the first 4 minutes during PA observations, while only 38% during MC. The distribution of the response times indicated a significantly higher frequency of affiliative interactions during the first two minutes of observation during PA than during MC (Figure 2; c2 = 4.5, p = .034, d.f. = 1). The mean response time during PA was 3.53 ± SD 2.74 min and 5.18 ± SD 2.88 min during MC, being this difference marginally significant (Wilcoxon paired test, z = 1.88, p = .059; n = 30).
The first affiliative interaction occurred earlier in 19 PA observations (attracted pairs) and in 11 MC sessions (dispersed pairs) than during the corresponding paired observation; 33 paired observations were neutral (no affiliative interaction in PC or MC). Among the 19 attracted pairs, the first affiliative interaction was initiated by the former receiver of the aggression in 12 cases (9 by females and 3 by males), and by the former aggressor in 7 cases (5 by females, 2 by males); however, there was no association between the role (aggressor-receiver) and the probability of initiating the affiliative interaction (c2 = 1.32, d.f. = 1; p = .25). With regards to the sex, females were significantly more prone to initiate the interaction (reconciliation) than males (c2 = 4.26, d.f. = 1, p = .039).
Figure 2.
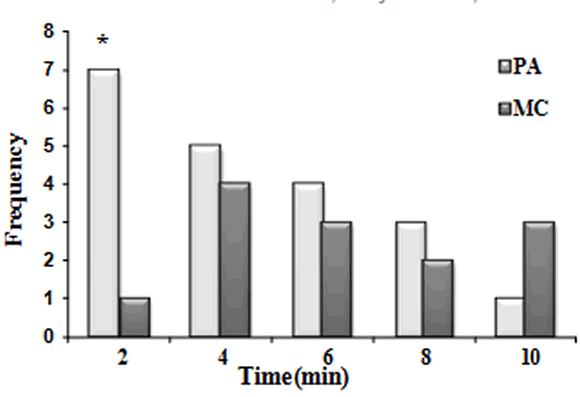
Distribution of response times during PA and MC sessions. A larger and significant number of interactions occur within the first 2 minutes of observation in the PA session than in the MC session (Goodness of fit c2 = 4.5, p = .034, d.f. = 1).
The overall CT of the group was 0.127 but with regards to the sex of the opponents, female-female interactions had a CT = 0.24 and female-male interactions a CT = 0.20. Male-male interactions resulted in dispersion between the individuals (CT = -0.07) and male-female ones never reconcile (CT = 0). The results were highly influenced by the interactions toward the third rank female (“Threadee”, Table 3), for which 57.1% of the agonistic interactions (4/7) resulted on dispersed pairs; these interactions were initiated by the alfa individuals (male and female) and the two subdominant males. In addition, this female never reconciled the agonistic interactions that she initiated (0/3) and only once did she receive reconciliation (1/7). When data from this female were removed from the analysis, the overall CT of the group was 0.196, female-female CT was 0.25, female-male CT was 0.42 and male-female CT was 0 again.
The Wilcoxon sign test performed by including all the data or by excluding the data from “Threadee” indicated that attracted pairs were marginally more probable than dispersed ones (Wilcoxon matched pairs test, all data, z = 2.5, p = .09; Mattracted = 2.11 ± SD 2.15; Mdispersed = 1.22 ± SD 1.48). Due to the small sample we could not analyze statistically the relation between dominance rank and conciliatory tendencies. However, qualitatively, there seem to be higher probability of reconciliation with rank (Table 3). The alfa male never received agonistic interactions, while the alfa female received two agonistic interactions from the top male and both were reconciled by her. The third-rank female and the alfa female reconciled the only agonistic interaction between them. The lower rank female received a high proportion of agonistic interactions but only 26% of them were reconciled (Table 3) and 66% resulted in neutral pairs (corresponding to 39% of all the neutral pairs in the group); an analogous pattern was found in the lower rank male. As a whole, qualitatively, the higher the rank of the individuals involved in an agonistic interaction, the higher the probability of attracted pairs and the lower the proportion of neutral pairs.
Table 3. Probability of reconciliation per dyad. The figures in parenthesis represent the number of attracted, neutral and dispersed pairs (PA-MC) respectively. Individuals are listed in their probable order of rank. The ranks below the top alfa male and female are not linear; the hierarchy of middle rank individuals (from Ringed Tail to Threadee) is not resolved.
4. Discussion
Agonistic interactions have a number of fitness-reducing consequences for the loser, such as missing the competed resource and increased likelihood of becoming the subject of renewed attacks either by the original aggressor or by third parties (references in Arnold & Aureli, 2007). In addition, agonism reduces the time spent in proximity between former opponents (Koyama, 2001), increases anxiety levels in aggressors and receivers (references in, Arnold & Aureli, 2007; Silk, 2002) and inter-individual distances (Arnold & Whiten, 2001; Pettit, Abegg, & Thierry, 1997); however, new evidences also emphasize an increase in the likelihood of affiliative interactions. In the present study, the probability of pacific reunions after an agonistic interaction was relatively low all together (12.7%), but it was higher in female-to- female (24-25%) and in female-to-male interactions (20-42%), this is because females have a higher probability of initiating affiliative interactions than males both after agonism and as whole. Interestingly, 50% of the PA-MC pairs were neutral, indicating that agonism did not turn out in dispersion nor affiliation (except for male-male interactions in which dispersion was the typical outcome), a result also found in other species of new world monkeys (Daniel et al., 2009; Leca et al., 2002; Manson et al., 2005; Perry, 1995); therefore, in general, the agonistic interactions did not have a disrupting effect in the dynamics of dyads. The conciliatory tendencies in our study group, global and per dyad type (by sex), were highly influenced by the behavior of the one subordinate female (“Threadee”), because she was involved in many agonistic interactions with subordinate males, which resulted in dispersed pairs. Therefore, the CT in this troop ranked from 0 (male-male interactions excluded) to 0.42 (female-male interactions, “Threadee” excluded), with overall conciliatory tendencies between 0.127 and 0.196.
Reconciliation in Cebidae has been analyzed in two species: the white-faced capuchins (Cebus capucinus, Leca et al., 2002; Manson et al., 2005; Perry, 1995) and the brown capuchin monkeys (Sapajus apella, Daniel et al., 2009; Verbeek & de Waal, 1997). In all these studies, the samples were taken from one or two groups only, and the number of PC-MC pairs used in the analysis ranged from 50 to 384 (see Daniel et al., 2009; Leca et al., 2002; Manson et al., 2005), thus, our pilot study falls within the sampling effort of capuchin monkeys’ studies allowing for substantial comparisons. Leca et al. (2002) examined 384 PC-MC pairs in a group of 12 individuals of white-faced capuchins, finding an overall CT of 0.237. The CT was higher among kin (PC-MC pairs = 54, CT = 0.481) than non-kin male-female dyads (PC-MC pairs = 340, CT = 0.212), thus suggesting that relatedness together with the value of the relationship are the most influential factors in determining the adaptive value of reconciliation in this species. On the other hand, Manson et al. (2005) found evidence of reconciliation in one of their two study periods with a free-ranging troop of C. capucinus (PC-MC pairs = 70, overall CT = 0.22; female-alpha male dyads CT = 0.44). In S. apella, Verbeek and de Waal (1997) demonstrated the occurrence of reconciliation (mean CT = 0.21) but only following fights that occurred in the absence of highly attractive food (agonistic interactions for food were not reconciled). Daniel et al. (2009) also found similar reconciliation tendencies in a captive group of 14 brown capuchins (PC-MC pairs= 190; overall CT = 0.298), which in addition, was more likely to occur between opponents that supported each other more frequently (“secure” fellows) and that spent more time together (“compatible” fellows) than in other dyadic relationships. Altogether, these evidences indicate relatively lower probabilities of reconciliation in Cebidae than in great apes (reviews in, Arnold & Aureli, 2007; Aureli, Cords, & van Schaik, 2002; Silk, 2002). The results of the present study correspond to the general tendency depicted from available information in Cebidae.
Several factors have been proposed to influence the probability of reconciliation in a troop, for instance: habit (arboreal vs. terrestrial), kinship, relationship quality, nature of the dispute (e.g., over food, refuge) and the type of social hierarchy (despotic vs. egalitarian) (references in Arnold & Aureli, 2007; Aureli et al., 2002; Silk, 2002). Among capuchins, kinship and relationship quality (Daniel et al., 2009; Leca et al., 2002; Manson et al., 2005), the age of the aggressor and the undecided outcome of the agonistic interaction (Daniel et al., 2009), non-food context (Verbeek & de Waal, 1997, but see Daniel et al., 2009 for a contrary result) and secure mother-offspring relationships (Weaver & de Waal, 2003) are positively related to an individual´s behavior during and after agonistic interactions and to the probability of reconciliation. Our study was not intended to determine the correlates of reconciliation, however, we have some data which partially support the relationship quality hypothesis (also known as valuable relationship hypothesis) (de Waal & Aureli, 1997). This hypothesis proposes that reconciliation restore, or at least improve, damaged high-quality relationships between former opponents (review in, Cords & Aureli, 2000). According to Cords and Aureli (2000) the quality of a relationship has three dimensions which are likely to influence the probability of reconciliation: value, compatibility and security. The relationship value (i.e., benefits derived from partnership) and partners´ compatibility (i.e., mutual tolerance and affiliation) are predicted to be positively associated with the likelihood of reconciliation whereas relationship security (i.e., probability of relationship change after a dispute) is predicted to be negatively associated, because insecure relationships are more likely than secure ones to be damaged by non-reconciled disputes. In primates, typically, female-male relationships have higher value as the rank of the male increases; female-female relationships are valuable and compatible, while male-male relationships have high value but low compatibility (Manson et al., 2005); therefore, reconciliation rate should decrease from female-male to male-male dyads. Our results indicate that female-female dyads and female-male dyads have the highest reconciliation rates, being even larger for female-male dyads when the subordinate female Threadee was not included in the analysis. Therefore, relationship quality could be important in the probability of reconciliation in Cebus olivaceus; howeverthis hypothesis is open to further investigation.
The balance between the importance of restoring damaged relationships and the risks involved in approaching a former opponent partially determines the probability of initiating reconciliation. The risks are thought to vary with the relative rank and the power asymmetry between former opponents as well as with the dominance style (egalitarian vs despotic) (Arnold & Aureli, 2007). Nonetheless, there is no general rule regarding the initiation of reconciliation. For instance, reconciliation is initiated by former aggressors in despotic (e.g., rhesus monkeys) and egalitarian species (e.g., bonobos) (references in Arnold & Aureli, 2007), and contrary to expected, by former victims in despotic long tailed macaques (Aureli, van Schaik, & van Hooff, 1989). In addition, it has been observed that individuals prone to initiate affiliative interactions ordinarily are also more prone to initiate reconciliation regardless of their role in the dispute. In our study, the probability of initiating reconciliation was independent of the role of the former opponents (aggressor or receiver) but dependent of the sex. As indicated before, females are more prone than males to initiate reconciliation, but also are more prone to affiliation in general (O’Brien, 1991). We presume that the lack of a bias regarding the initiation of reconciliation by aggressors or receivers in the present study is due to: a) the poorly developed hierarchy below the top male and female, b) the relatively high proportion of agonistic interactions involving subordinates, both as aggressor and as victims, c) the tolerant temperament of the species (Fragaszy et al., 2004), and/or d) the proclivity of females to engage in affiliative behaviors in general (O´Brien, 1991). All of these possibilities remain to be tested in long-term studies.
Recently, several authors have stressed the limitations of the variables traditionally used to measure reconciliation (e.g., the response time) and the need for the inclusion of other variables such as the frequency and duration of affiliations and latencies to second and third affiliative interactions after an agonistic interaction (Logan et al., 2013; Manson et al., 2005). Logan et al. reasoned that the second and third affiliative interaction may not simply be a response to the previous one, but a combined response to the previous agonistic interaction. Interestingly, in our study, secondary affiliations were observed during PA period and also during MC, thus repeated affiliation does not seem to relate to the previous agonism in C. olivaceus. In the present study, we also analyzed the duration and frequency of affiliations, both in PA and MC observation sessions. These variables however, did not differ significantly between sessions and the traditional methods were more informative; we yet propose to include them in any study of reconciliation because significant differences have been found in other species and deserve to be further explored (e.g., Logan et al., 2013). We also propose that the length of the observation period must be flexible and adjustable with regards to the habits of the study species, and should not be limited to the registration of the first affiliative interaction. For instance, Manson et al. (2005) found that the probability of renewed aggression decreased as the first PC affiliation was delayed after the initial conflict in C. capucinus; therefore a premature affiliation does not necessarily indicates actual reconciliation, and delayed affiliation must be most relevant(see Rolland & Roeder, 2000).
The habit of the species and the possibility to move fast over long distances may also have an influence on the response time and stresses the need of measuring more dependent variables in reconciliation studies, as indicated previously. For instance, in captive C. capucinus most reconciliation occurred within the first 2 min after an agonistic interaction (Leca et al., 2002), much as in other species (Kappeler & van Schaik, 1992); however, in the wild, reconciliation appears to be significantly delayed, occurring later than30 min after the end of a agonistic interaction (Leca et al., 2002; Perry, 1995). In general, wild arboreal monkeys maintain large inter individual distances while foraging (Fragaszy et al., 2004; Phillips, 1995; Robinson, 1988) and usually rest afterwards, thus the opportunity for reconciliations is postponed until the group reunites again (Verbeek & de Waal, 1997). This condition seems to be applicable to C. capucinus (Leca et al., 2002) and S. apella (Manson et al., 2005; Verbeek & de Waal, 1997) but it is not supported by findings in other highly arboreal monkeys, such as long-tailed macaques (Aureli, 1992; Aureli et al., 1989).
Finally, from the low conciliatory tendencies found in the three capuchins studied to date, we propose that reconciliation may not be a fundamental mechanism in maintaining social cohesion in Cebidae. Given that a large proportion of agonistic interactions result in neutral pairs, it is likely that most of them do not have a damaging effect in dyadic relationships. Therefore, only those agonistic interactions with a high potential to damage a relationships would be reconciled (Daniel et al., 2009). The habits of the species must also have an important effect both on the disrupting effect of agonistic interactions and on the probability of reconciliation which needs to be investigated.
Acknowledgements
We are indebted to J. Rodríguez, R. A. Rojas, and N. Marín for authorizing us to perform the observations at Parque Generalísimo Francisco de Miranda, and to A. Quintero and the animals´ keepers for their support while conducting the study. Z. Tárano wishes to thank J. Medina for revising the English version of the manuscript.
References
Alexander, R. D. (1974). The evolution of social behaviour. Annual Review of Ecology and Systematics, 5, 325-383. doi: 10.1146/annurev.es.05.110174.001545
Altman, [ Links ] J. (1974). Observational study of behavior: Sampling methods. Behaviour, 49, 227-265.doi: 10.1163/156853974X00534
Arnold, [ Links ] K., & Aureli, F. (2007). Post-conflict reconciliation. In J. Campbell, A. Fuentes, K. C. MacKinnon, M. Panger, & S. K. Bearder, (Eds.), Primates in Perspective (pp. 592-608). Oxford: Oxford University Press. [ Links ]
Arnold, K., & Whiten, A. (2001). Post-conflict behaviour of wild chimpanzees (Pan troglodytes schweinfurthii) in the Budongo Forest, Uganda, Behaviour, 138, 649-690. doi: 10.1163/156853901316924520
Aureli, [ Links ] F. (1992). Post-conflict behaviour among wild long-tailed macaques (Macaca fascicularis). Behavioural Ecology and Sociobiology, 31, 329-337. doi: 10.1007/BF00177773
Aureli, [ Links ] F., Cords, M., & van Schaik, C. P. (2002). Conflict resolution following aggression in gregarious animals: A predictive framework. Animal Behaviour, 64, 325-343. doi: 10.1006/anbe.2002.3071
Aureli, [ Links ] F., van Schaik, C. P., & van Hooff, J. A. R. A. M. (1989). Functional aspects of reconciliation among captive long-tailed macaques (Macaca fascicularis). American Journal of Primatology, 19, 39-51. doi: 10.1002/ajp.1350190105
Cools, [ Links ] A. K. A., van Hout, A. J. M., & Nelissen, M. H. J. (2008). Canine reconciliation and third-party-initiated post-conflict affiliation: Do peacemaking social mechanisms in dogs rival those of higher primates? Ethology, 114, 53-63. doi: 10.1111/j.1439-0310.2007.01443.x
Cords, [ Links ] M., & Aureli, F. (2000). Reconciliation and relationship qualities. In F. Aureli, & F. B. M. de Waal (Eds.), Natural Conflict Resolution (pp. 177-198). Berkeley & Los Angeles CA: University of California Press
Daniel, [ Links ] J. R., Santos, A. J., & Cruz, M. G. (2009). Post-conflict behaviour in brown capuchin monkeys (Cebus apella). Folia Primatologica, 80, 329-340. doi: 10.1159/000258647
Eisenberg, [ Links ] J., & Redford, K. (1989). Mammals of the Neotropics: The Northern Neotropics. Chicago: University of Chicago Press. [ Links ]
Fragaszy, D., Visalberghi, E., & Fedigan, L. (2004). The complete Capuchin: The Biology of the Genus Cebus. Cambridge: Cambridge University Press. [ Links ]
van Hooff, J. A. R. A. M., & van Schaik, C. P. (1992). Cooperation in competition: The ecology of primate bonds. In A. H. Harcourt, & F. B. M. de Waal (Eds.), Coalitions and Alliances in Humans and Other Animals (pp. 357-389). Oxford: Oxford University Press. [ Links ]
Kappeler, P. M. (1993). Reconciliation and post-conflict behaviour in ringtailed lemurs, Lemur catta, and redfronted lemurs, Eulemur fulvus rufus. Animal Behaviour, 45, 901-915. doi: 10.1006/anbe.1993.1110
Kappeler, [ Links ] P. M., & van Schaik, C. P. (1992). Methodological and evolutionary aspects of reconciliation among primates. Ethology, 92, 51-69. doi: 10.1111/j.1439-0310.1992. tb00948.x
Koyama, [ Links ] N. F. (2001). The long-term effects of reconciliation in Japanese macaques Macaca fuscata. Ethology, 107, 975-987. doi: 10.1046/j.1439-0310.2001.00731.x
Leca, [ Links ] J. B., Fornasieri, I., & Petit, O. (2002). Aggression and reconciliation in Cebus capucinus. International Journal of Primatology, 23, 979-998. doi: 10.1023/A:019641830918
Logan, [ Links ] C. J., Emery, N. J., & Clayton, N. S. (2013). Alternative behavioral measures of post-conflict affiliation. Behavioral Ecology, 24, 98-112. doi: 10.1093/beheco/ars140
Ló [ Links ]pez, M. CH., & Tárano, Z. (2008). Comportamiento social del mono capuchino común Cebus olivaceus (Primates: Cebidae) en tres exhibiciones zoológicas de Caracas, Venezuela. Revista de Biología Tropical, 56, 1503-1520. [ Links ]
Manson, J. H., Perry, S., & Stahl, D. (2005). Reconciliation in wild white-faced capuchins (Cebus capucinus). American Journal of Primatology, 65, 205-219.doi: 10.1002/ajp.20110
Nieuwenhuijsen, [ Links ] K., & de Waal, F. B. M. (1982). Effects of spatial crowding on social behavior in a Chimpanzee colony. Zoo Biology, 1, 5-28. doi: 10.1002/zoo.1430010103
O’ [ Links ]Brien, T. G. (1991). Female-male social interactions in wedge-capped capuchin monkeys: benefits and costs of group living. Animal Behaviour, 41, 555-567.doi: 10.1016/S0003-3472(05)80896-6
Palagi, [ Links ] E., Antonacci, D., & Norscia, I. (2008). Peacemaking on treetops: First evidence of reconciliation from a wild prosimian (Propithecus verreauxi). Animal Behaviour, 76, 737-747. doi: 10.1016/j.anbehav.2008.04.016
Perry, [ Links ] S. (1995). Social relationships in wild white-faced capuchin monkeys, Cebus capucinus (Unpublished Doctoral Dissertation). University of Michigan, USA. [ Links ]
Petit, O., Abegg, C., & Thierry, B. (1997). A comparative study of aggression and conciliation in three cercopithecine monkeys (Macaca fuscata, Macaca nigra, Papio papio), Behaviour, 134, 415-432. doi: 10.1163/156853997X00610
Phillips, [ Links ] K. (1995). Resource patch size and flexible foraging in white-faced capuchins (Cebus capucinus). International Journal of Primatology, 16, 509-519. doi: 10.1007/BF02735800
Polizzi di Sorrentino, E., Schino, G., Visalberghi, E., & Aureli, F. (2010). What time is it? Coping with expected feeding time in capuchin monkeys. Animal Behaviour, 80, 117-123. doi: 10.1016/j.anbehav.2010.04.008
Robinson, [ Links ] J. G. (1981). Spatial structure in foraging groups of wedge-capped capuchin monkeys Cebus nigrivittatus. Animal Behaviour, 29, 1036-1056. doi: 10.1016/S0003-3472(81)80057-7
Robinson, [ Links ] J. G. (1988). Demography and group structure in wedge-capped capuchin monkeys, Cebus olivaceus. Behaviour, 104, 202-231. doi: 10.1163/156853988X00520
Rolland, [ Links ] N., & Roeder, J. J. (2000). Do ringtailed lemurs (Lemur catta) reconcile in the hour post- conflict?: A pilot study. Primates, 41, 223-227. doi: 10.1007/BF02557804
Silk, [ Links ] J. B. (2002). The form and function of reconciliation in primates. Annual Review of Antropology, 31, 21-44. doi: 10.1146/annurev.anthro.31.032902.101743
Smuts, [ Links ] B. B., Cheney, D. L., Seyfarth, R. M., & Wrangham, R. W. (1986). Primate Societies. Chicago: University of Chicago Press.
Sokal, R. R., & Rohlf, F. J. (1995). Biometry: The Principles and Practice of Statistics in Biological Research. New York: W. H. Freeman and Company. [ Links ]
Tárano, Z. & López, M. CH. (2015). Behavioural repertoires and time budgets of semi-captive and captive groups of the wedge-capped capuchin monkey Cebus olivaceus. Folia Primatológica, 86, 203-222. doi: 10.1159/000381397
Thierry, [ Links ] B., Aureli, F., Nunn, C., Pettit, O., Abegg, C., & de Waal, F. B. M. (2008). A comparative study of conflict resolution in macaques: insights into the nature of trait covariation. Animal Behaviour, 75, 847-860. doi: 10.1016/j.anbehav.2007.07.006
Veenema, [ Links ] H. C., Das, M., & Aureli, F. (1994). Methodological improvements for the study of reconciliation. Behavioural Processes, 31, 29-38. doi: 10.1016/0376-6357(94)90035-3
Verbeek, [ Links ] P., & de Waal, F. B. M. (1997). Post-conflict behaviour of captive brown capuchins in the presence and absence of attractive food. International Journal of Primatology, 18, 703-725. doi: 10.1023/A:1026391728909
Visalberghi, [ Links ] E., & Fragaszy, D. (1995). The behaviour of capuchin monkeys, Cebus apella, with novel food: The role of social context. Animal Behaviour, 49, 1089-1095. doi: 10.1006/anbe.1995.0137
de Waal, [ Links ] F. B. M. (2000). Primates - A natural heritage of conflict resolution. Science, 289, 86-590. doi: 10.1126/science.289.5479.586
de Waal, [ Links ] F. B. M., & Aureli, F. (1997). Conflict resolution and distress alleviation in monkeys and apes. In C. S. Carter, B. Kirkpatrick, I. Lenderhendler (Eds.), The Integrative Neurobiology of Affiliation (pp. 317-328). New York: Annals of the New York Academy of Sciences.
de Waal, F. B. M., & van Roosmalen, A. (1979). Reconciliation and consolation among chimpanzees. Behavioural Ecology and Sociobiology, 5, 55-66. doi: 10.1007/BF00302695
de Waal, [ Links ] F. B. M., & Yoshihara, D. (1983). Reconciliation and redirected affection in rhesus monkeys. Behaviour, 85, 224-241. doi: 10.1163/156853983X00237
Weaver, [ Links ] A., & de Waal, F. B. M. (2003). The mother-offspring relationship as a template in social development: Reconciliation in captive brown capuchins (Cebus apella). Journal of Comparative Psychology, 117, 101-110. doi: 10.1037/0735-7036.117.1.101
Welker, [ Links ] C., Becker, P., Höhmann, H., & Schäfer-Witt, C. (1987). Social relations in groups of the black-capped capuchin Cebus apella in captivity. Interactions of group-born infants during their first six months of life. Folia Primatologica, 49, 33-47. doi: 10.1159/000156306 [ Links ]
Citar este artículo como: Tárano, Z. y Flores, R. M. (2016). Analysis of post-agonism reconciliation in a captive group of the wedge-capped capuchin monkey, Cebus olivaceus: a pilot study. Revista Argentina de Ciencias del Comportamiento, 8(1), 50-60

