











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCTION
Asplenium L. is taxonomically one of the most complex fern genera with about 700 species, nearly cosmopolitan in distribution (PPG I, 2016; Xu et al., 2020). It is characterized by usually having erect rhizomes with clathrate scales and dictyostelic or radial steles, small to medium-sized leaves that are simple to 4-pinnate, laminae with nonarticulate petioles and distinctly sulcate rachises with a raised ridge in the centre and two grooves on each side, rachis-costa architecture characterized by a terete and alate rachis with wings that are confluent with the basiscopic pinna margin, usually free veins, elongate sori with a narrow indusium borne on a vein, and a base chromosome number x = 36 (Murakami & Moran, 1993; Cheng & Murakami, 1998; Lin & Viane, 2013; Sundue & Rothfels, 2014; Xu et al., 2020).
In Argentina, Asplenium constitutes the most species-rich fern genus, with 38 taxa assigned to the native flora, distributed mainly in the Neotropical area of the country, with few species in the Andean region (Ganem et al., 2016; Arana et al., 2020). Among the Neotropical species, Asplenium achalense Hieron. was originally described by Hieronymus (1896) based on three specimens collected in Pampa deAchala, Córdoba province, central Argentina. Previously, Grisebach (1874, 1879) misapplied the name Asplenium furcatum Thunb. (an African species), to the specimens of Asplenium achalense collected by C. Lorentz. This mistake was corrected later by Hieronymus, who described the species as a new taxon.
Traditionally, A. achalense was regarded as an endemic species of Argentina related to A. serra Langsd. & Fisch., which was also cited for the Argentine flora (de la Sota, 1977; Arana et al., 2004; Ganem et al., 2007, 2016). However, some authors have considered that the differences between A. achalense and A. serra remain poorly understood, suggesting even that the latter could be a variant of A. achalense (Kessler & Smith, 2006, 2018). Asplenium achalense has also been regarded as a complex of species that includes A. incurvatum Fée, A. micropaleatum M. Kessler & A. R. Sm., A. mosetenense M. Kessler, and A. tunquiniense M. Kessler & A. R. Sm., all from Bolivia, and distinguished only by morphological differences of the rhizomatic scales (Kessler & Smith, 2006, 2018; Jonas, 2011). Analysing the specimens deposited in different herbaria of the world, only a few were correctly identified as Asplenium achalense, the first collections date back to the 19th century and the latest are from three decades ago, at the end of the 20th century. This species was not longer found in the “locus classicus” (Pampa de Achala, Córdoba province) after several field trips from 1999 to 2022. Recently, in August 2022, we located a population of A. achalense in the proximities of the locality of Cochuna, Tucumán province, which allowed us to analyse both, morphological characters in living material and the species habitat.
Accurate taxonomic identification is essential to estimate the biodiversity of an area and constitutes the basis to carry out ecological and biogeographical studies, and to propose conservation actions. Therefore, the aim of this work is to provide a detailed analysis of the diagnostic morphological and anatomical features of Asplenium achalense sporophyte, including spore morphology, as a contribution to clarify the taxonomic controversies about this taxon. Also, information on its distribution area and the habitats where it grows is provided.
MATERIALS & METHODS
Field observations were carried out at Cochuna, Tucumán province, during winter 2022. Photographs of the habitat and the plant were taken “in situ” (Fig. 1A-B). The specimens were deposited in herbarium RCVC (acronym according to Thiers, 2022). Herborized specimens housed at herbaria of Fundación Miguel Lillo (LIL), Universidad Nacional de Córdoba (CORD), Museo de La Plata, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata (LP), and Instituto de Botánica Darwinion (SI) were also employed in this study. In addition, high resolution digital images of specimens from B and MO were analysed.
Morphological and anatomical studies were performed under a stereoscopic microscope Nikon SMZ1000 and a light microscope (LM) Nikon E200. Pinnae were diaphanized according to the Foster’s technique, stained with safranin and mounted in jelly- glycerin (D’Ambroggio de Argüeso, 1986). Rhizome and leaf scales, as well as spores, were mounted in 20% glycerin without prior treatment. The characters analysed were: degree of laminae division, venation pattern, indument of leaves and rhizome, location of sori, indusium and spore morphology.

Fig. 1 Habitat and living specimen of Asplenium achalense. A, montane forest of Yungas, Tucumán province. B, epiphytic habit. Color version at http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/1080/1276
For scanning electron microscope (SEM), the spores were mounted on aluminium stubs also without pre-treatment and covered with gold for 10 minutes. Observations were made in a microscope FEI Quanta 200 model (Servicio de Microscopía Electrónica de Barrido y Microanálisis del LIMF, Facultad de Ingeniería, Universidad Nacional de La Plata) with a 15 kV voltage. For descriptions of the sculpture of the spore wall with SEM, as well as tha anatomy of the petiole and root, the works of Giudice et al. (2002), Ganem et al. (2019) and Luna et al. (2020) were consulted. The microscopic slides used in this study are stored in the “Laboratorio de Anatomía Comparada, Propagación y Conservación de Embriofitas “Dr. Elías R. de la Sota”, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata.
RESULTS
Asplenium achalense Hieron., Bot. Jahbr. 22: 378. 1896. Type: Argentina. Córdoba. Depto. Punilla: Sierra Achala, entre Tanti y el Río Juspe, 12-II-1876, G. Hieronymus 846 (lectotype B-20-009946 digital image! designated by Cremers & Viane, Syst. Geogr. Plants 78(2): 218. 2008; isolectotypes: B-20-009945 digital image!, B-20-009947 digital image!, CORD 00001377!, SI!). Fig. 2.
Root anatomy. The root structure of A. achalense is described in detail in the work of Luna et al. (2020). Briefly, the most relevant anatomical characteristic of Asplenium roots is the presence of “asplenium sclereids” in the inner cortex. In the case of A. achalense, the inner cortex is made up by a single layer of six sclereids, which corresponds to the Aethiopicum root subtype described by Schneider (1997) and proposed later by Schneider et al. (2004) as the synapomorphy of Asplenium section Sphenopteris Mett. (Luna et al., 2020).
Rhizome. Compact to short creeping, 3-7 mm in diameter (excluding scales); rhizome scales lanceolate to linear-lanceolate, clathrate, 3-5 mm long, bright dark brown to blackish, margin entire or with 1-2 teeth near the basis; apices long attenuate, minutely remotely denticulate (Fig. 3A-C).
Leaves. Pendant, up to 80 cm (Fig. 1); petioles 25-40 cm × 2-4 mm, ca. 1/3 to 1/2 of the leaf length, dull brown, sulcate adaxially. Petiole bases scaly, scales similar to those of the rhizome, glabrescent distally, sparsely scaly, the scales linear to subulate; and with scarcely glandular-pubescent near the lamina basis, hairs bicellular with apical rounded cell (Fig. 3D-H).
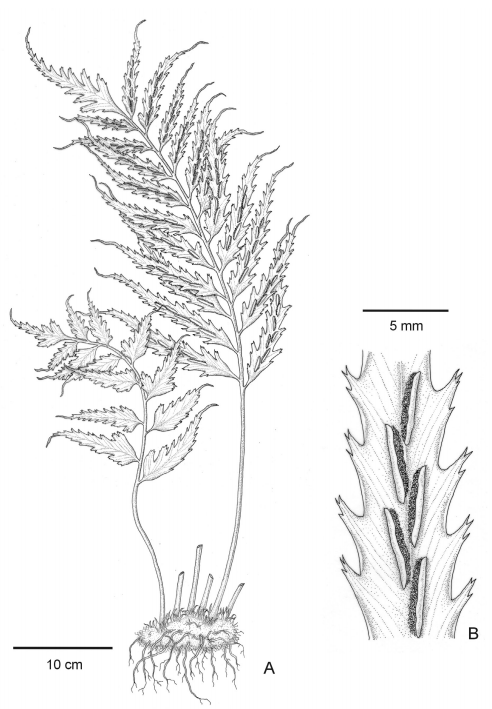
Fig. 2 Asplenium achalense. A, entire specimen with a juvenile leaf (left) and an adult leaf (right). B, detail of pinna margin and sori.

Fig. 3 Indument of Asplenium achalense sporophyte. A, rhizome covered by scales. B, micrograph of rhizomatic scales under stereoscopic microscope. C, LM micrograph of a scale where the thickened radial cell walls and smooth margins are observed in more detail. D, base of petiole with scales. E, bicellular hairs of petiole (arrows). F, detail of a hair under LM. G-H, scales present in the middle portion of the petiole, which are intermingled with the hairs. G, larger scales. H, smaller scales. I, scales in the rachis. J, LM micrograph of a scale. K, leaf scales (arrows). L, detail of a scale under LM. Abbreviations: h, hair; lp, leaf primordium; ls, leaf scale; rhi, rhizome; so, sorus. Color version at http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/ view/1080/1276
Petiole anatomy: the structure of the petiole is described in the publication of Ganem et al. (2019). The main characteristics observed in cross section are the presence of disrupted subepidermal mechanical tissue, absence of circumendodermal band, presence of two lateral aeration lines and x-shaped contour of the xylem in the middle zone.
Blades: once-pinnate to pinnate-pinnatifid at the bases, broadly lanceolate, widest at the bases, the apices acuminate, conform, non-proliferous, the bases truncate. Rachises adaxially grooved, dark brown coloured, with scattered, subulate brown scales 0.7-1.2 mm long; sparsely glandular pubescent, hairs similar to those on the petioles (Fig. 3I-J).
Pinnae: 10-20 pairs, short-petiolulate (1-5 mm), petiole green coloured to the costal bases; subcoriaceous, almost glabrous on both sides, abaxially with scattered brown linear to subulate scales similar to those of the rachis (Fig. 3K-L), especially near the costae; pinnae lanceolate to linear lanceolate, the bases cuneate to truncate, apices long- acuminate, up to 110 mm × 10 mm, the basal ones sometimes auriculate, margin strongly biserrate, veins 1-2 forked, each secondary vein ending in a marginal teeth. Apical pinnae pinnatifid (Fig. 1B, Fig. 2).
Sori: linear, 15-28 per pinna, placed on lowermost acroscopic vein branch on both sides, close to the midvein and nearly parallel to it; indusia ca. 4-12 × 0.8-1 mm, light brown, glabrous, entire (Fig. 2B, Fig. 3K, Fig. 4A).
Spores. The description and measurements of the spores and the spore wall are given in Giudice et al. (2002). The spores are monolete, brownish under LM, 39 × 36 µm in equatorial diameter. Perispore with fused ridged folds with a smooth margin. The surfaces between the folds are rugulated and scarcely perforated (Fig 4B-C).
Distribution and habitat. This species is known only from north-western and central Argentina, originally found growing as saxicolous in highland grasslands at 1200-2000 m a.s.l. These environments belong to the Comechingones biogeographic province in central Argentina (Arana et al., 2017, 2021). Later, during the 20th century, the species was found as epiphyte in humid montane forests of Tucumán and Salta provinces, in habitats of the Yungas biogeographic province (de la Sota, 1977).
After several trips during 1999 to 2022 covering the extension of Comechingones and Yungas in Argentina, the species was only found in the last environment, growing epiphytically at the uppermost part of the Blepharocalyx salicifolius (Kunth) O. Berg (Myrtaceae), a typical species from Yungas (Fig. 1). Previously, A. achalense was mentioned as epiphyte on Ocotea porphyria (Griseb.) van der Werff (Lauraceae), an endemic species of Montane Forest district of Yungas (Arana et al., 2021).
Additional specimens examined
ARGENTINA. Córdoba . Depto. Punilla: Sierra de Achala, Río Juspe; Cuesta de Copina, 19-II-1877, Hieronymus s.n. (B-20-009949, CORD); Sierra Achala, entre Tanti Cuchi del Rosario y el Río Juspe, 7-I-1881, Hieronymus s. n. (B-20 009948, CORD). Tanti, II-1897, Stuckert 2261 (SI). Ollada cerca de la Pampa de San Luis, X-1874, Lorentz s.n. (CORD, LP). Casas Viejas, entre Olaen y Pampa de San Luis, 21- II-1899, Kurtz 10539 (CORD). Los Mogotes, Capilla del Monte, 01-II-1917, Hosseus 235 (CORD). Sierra Chica, Las Higueras, in fisuris umbrosis rupium, XII-1875, Kurtz 2843 (SI). Depto. San Alberto: Mina Clavero, 12-XII-1901, Stuckert 10538 (CORD). Salta. Depto. Anta: Parque Nacional El Rey, camino a Pozo verde, pasando el primer cruce, epífito sobre laurel, 19- XI-1981, Brown 1622 (MO, SI). Depto. La Caldera: Camino de cornisa, Alto La Sierra, 8-XI-1988, Palaci 1036 (MCNS). Depto. Santa Victoria: 2438,51 m s.m., 20-VIII-1993, Hilgert 755 (SI). Tucumán. Depto. Famaillá: Caspichango, VII-1912, Castillón s. n. (SI 16141). Depto. Monteros: quebrada a Caspichango, epífita, 800 m s.m., 4-X-1929, Venturi 9585 (SI). Quebrada a Caspichango, epífita, adherente a la corteza, 16-II-1907, Dinelli 668 (SI). Quebrada Pueblo Viejo, 1-I-1965, de la Sota 4060 (LP). Cochuna, epífito sobre horco molle, 10-VIII-2022, Arana 2053 (RCVC). Depto. Tafí: Quebrada del Tafí, epífita, 1500-2000 m s.m., VII-1920, Venturi 879 (SI).
DISCUSSION AND CONCLUSIONS
Reticulate evolution is a common mechanism in fern evolution, reported in the genus Asplenium whose numerous taxa can form species complexes with a morphological continuity between them (Dyer et al., 2012; Schneider et al., 2013; Liang et al., 2021).
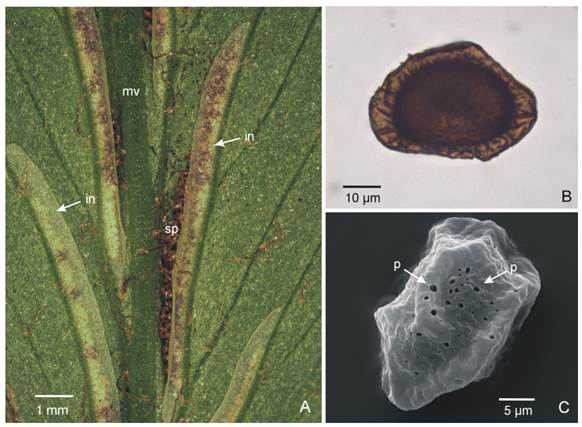
Fig. 4 Sori and spores of Asplenium achalense. A, abaxial surface of a pinnae showing the elongated indusia covering the sporangia (arrows). B, LM micrograph of a spore where the brownish coloration can be observed. C, SEM micrograph of a spore showing the perispore in detail. Arrows point to perforations. Abbreviations: in, indusium; mv, midvein; p, perforations; sp, sporangia. Color version at http://www.ojs.darwin.edu.ar/index.php/darwiniana/article/view/1080/1276
The Asplenium species complex surrounding A. serra, as delimited by Tryon & Stolze (1993), is highly variable in several characteristics as the rhizome scale colour and size; pinna size, number and margin, and degree of pinna dissection (Kessler & Smith, 2006). Particularly, the distinction between A. achalense and A. serra, as well as among related species (A. incurvatum, A. micropaleatum, A. mosetenense, and A. tunquiniense), remain poorly understood (Kessler & Smith, 2006, 2018).
According to Jonas (2011), distinctions between A. achalense and A. serra are supported by a molecular and morphometric data. However, the Bolivian species A. incurvatum, A. mosetenense, and A. tunquiniense recognized by Kessler & Smith (2006) were found by Jonas (2011) to nest within A. achalense, using both molecular and morphometric data, and differentiated only by rhizome scales features (Kessler & Smith, 2018). After the analyses of both, living material and herborized specimens from Argentina, along with the observation of digital images of specimens from Bolivia (erroneously referred to A. achalense), we presume that the Bolivian materials used by Jonas (2011) for the molecular analyses do not correspond to the true A. achalense, but to A. serra or other unnamed related species of this complicated species complex.
By other hand, in classical Argentinean floras (de la Sota, 1977; Ganem et al., 2007; 2016) the specimens referred to “Asplenium serra” are not identical to the taxon A. serra as delimited by Kessler & Smith (2006), so we consider that they could belong to an unnamed species so far, erroneously called A. achalense by Kessler & Smith (2006, 2018) and Jonas (2011). In this way, it is probable that the true A. serra does not occur in north-western Argentina (Jonas, 2011), although we provisionally chose to keep this name until the ongoing studies are completed. As the type of A. serra was collected in south-eastern Brazil, elucidation of the identity of the specimens referred to this taxon in Argentina requires an exhaustive analysis of specimens from Bolivia, south-east Brazil and north-western Argentina, by comparing the protologues and the type specimens, inquiring into diagnostic morphological and anatomical features, including indument and spore morphology, to accurately differentiate the taxa. All these studies are currently being conducted by this research group.
Also, the juvenile plants attributed to A. achalense from Bolivia have strongly dissected leaves that are often up to 1-pinnate-pinnatisect in earliest leaves, and become gradually less dissected as plants grow (Kessler & Smith, 2018). According to our observations, the juvenile plants of the true A. achalense have pinnae resembling those of the mature plants but wider. Only with the increase in age and size the leaves tend to have pinnae with more strongly reduced blade, which become linear lanceolate at maturity (i.e. with sori).
The taxonomic importance of spore morphology and wall sculpture in lycophytes and ferns is generally well established; spore morphology using scanning electron microscopy is very useful in the taxonomy of complex ferns taxa (e.g. Braggins & Large, 1990; Vaganov et al., 2017; Adeonipekun et al., 2021; Gorrer et al., 2021; Morajkar et al., 2021; Irfan et al., 2022). Particularly, Asplenium spores are variable in size, shape and perispore morphology and ornamentation, being consistently monolete, frequently ellipsoidal, having prominent folds with alate, costate, cristate, echinate, and reticulate perispore which is usually fenestrated in areolae between the folds (Regalado & Sanchez, 2002; Ganem et al., 2013; Shah et al., 2020; Irfan et al., 2022). Concerning spore morphology and wall sculpture of A. achalense and related species, Viane (1992) mentioned that A. achalense and A. serra share the same reticulate perispore pattern. In the present work, we found remarkable differences in spore wall sculpture patterns that allow differentiating the species by themselves. In contrast to what was described for the spores of true Asplenium achalense, the spores of Argentinean “A. serra” are 48 × 45 µm in equatorial diameter, the perispore is reticulated, has winged folds with equinulated margins and large and numerous perforations (Giudice et al., 2002), similar to the spores of A. serra described by Tryon & Lugardon (1991) using Dominican Republic specimens, which belong to Hispaniola biogeographic province, Antillean subregion of Neotropical region (Morrone et al., 2022).
According to Kessler & Smith (2018), “Asplenium achalense” may best be interpreted as a species complex. In this contribution we provide valuable diagnostic features present in rachises and laminae scales, along with characters as glandular hairs present in petioles and rachises, and spore wall features, that are critical to delimitate the true Asplenium achalense as a well-defined taxon, endemics of north-west to central Argentina. Remains pending further research to elucidate the identity of the Argentinean specimens regarded as “A. serra” and Bolivian specimens referred to “A. achalense”.
Key to the species of Asplenium serra complex present in Yungas biogeographic province
1. Blades 1-pinnate-pinnatisect to 2-pinnate-pinnatifid, at least proximal pinnae incised fully to the costae .... A. tunquiniense
1. Blades 1-pinnate, if proximal pinnae incised, then not fully to the costae ................................................................. 2
2(1). Rhizome scales up to 2 mm long ................................................................................................. A. micropaleatum
2. Rhizome scales 3-18 mm long .................................................................................................................................... 3
3(2). Rhizome scales 6-18 mm long ............................................................................................................................... 4
3. Rhizome scales 3-5 mm long ...................................................................................................................................... 5
4(3). Rhizome scales dark brown to blackish ............................................................................................. A. incurvatum
4. Rhizome scales orange to castaneous ................................................................................................... A. mosetenense
5(4). Petioles and rachises scaly and glandular-pubescent, hairs bicellular with apical rounded cell. Pinnae margin strongly biserrate. Apical pinnae pinnatifid. Spores 39 × 36 µm, perispore folded- ridged, fused ridged folds with a smooth margin and scarcely perforated ........................................................................................................ A. achalense
5. Petioles and rachises only scaly, without such hairs. Pinnae margin only slightly serrate. Apical pinnae conform. Spore 48 ×
45 µm, perispore folded- winged, winged folds with equinulated margins and large and numerous perforations ....“A. serra”

