











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
The Ediacara Biota, named after the Ediacaran Hills in the Flinders Ranges region of South Australia, were among the first multicellular organisms to inhabit the shallow coastal waters of our planet, some 555 million years ago. By studying the first emergence of complex creatures, with an assortment of shapes, sizes, and symmetries, we gain insights into the underlying natural processes behind the rise and evolution of early multicellular life and the ecosystems they were a part of. Phyllo%oon hanseni was originally described by Jenkins and Gehling (1978) from three specimens collected at Devil’s Peak, south of Quorn, in the Flinders Ranges of South Australia (Fossil site 3, Fig. 1A), and belongs to the White Sea Assemblage of the Ediacara Biota (Droser et al, 2017). Superficially, P. hanseni is frond-like in appearance, with a peculiar type of symmetry called ‘glide reflection symmetry’, where the two halves are roughly similar in shape and size but are slightly offset longitudinally (see Fig. 2). These two halves are made up of repeating long, rounded units, each based in a groove in a central corrugation that extends the length of the organism. Specimens range in length from as little as 45 mm to well over 300 mm, up to 60 mm in width, and can grow more than 125 units per side with many articulating interrupted off broken slab edges. P. hanseni also stands out due to its bizarre mode of preservation, in which it leaves positive hyporelief impressions, rather than the negative hyporelief preservation typical of many other fossils in this assemblage, such as Spriggina, Dickinsonia or Tribrachidium (Jenkins and Gehling, 1978). This has led some to suggest that this was a delicate, soft-bodied organism, made up of quilted, air-mattress like units, which quickly burst and decay after burial, but before the overlying sediment hardened (Gehling, 1999). It is known exclusively from the wave-base sands and sheet-flow sands facies, alongside two other taxa with the same preservation mode, Aulozoon’ and Funisia (Reid et al., 2020). Some have speculated that P. hanseni was a grazing trace fossil (Ivantsov, 2013), and it has even been suggested that it may be part of a terrestrial biota (Retallack, 2013). This last claim is widely disputed, as local sedimentary features imply large scale marine inundation at this time (Tarhan et al., 2017). This report hopes to uncover any trends in morphological parameters of Phyllozoon and will attempt an accurate description of this organism. We will also briefly discuss the palaeoenvironment, but purely within the biological context of P. hanseni, as an in-depth investigation into the geological nature of this deposit has been addressed by other authors (Gehling and Droser, 2012).
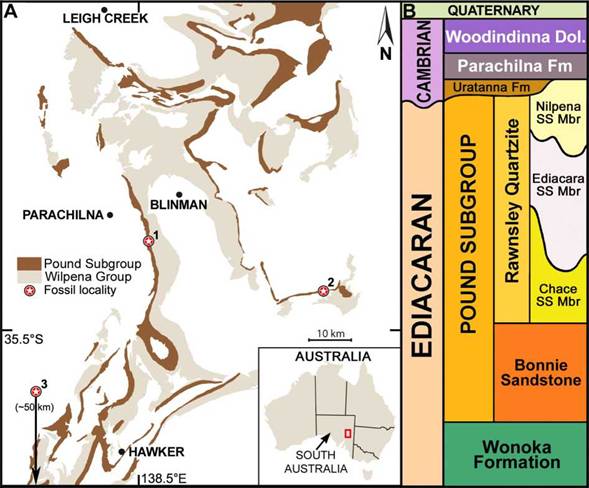
Figura 1: A) Mapa geológico de la Sierra de Flinders, Australia del Sur (Modificado de Reid et al.. 2020). Localidades con especímenes de Phyllozoon hanseni marcados con estrellas numeradas 1, 2 y 3, que representan Bathtub Gorge, Tooth Knob y Devil’s Peak respectivamente; B) Diagrama que muestra la sucesión estratigráfica simplificada en la Sierra de Flinders. /Figure 1. A) Geological map of the Flinders Ranges, South Australia (Modified from Reid et al 2020). Sites bearing Phyllozoon hanseni marked by numbered stars 1, 2 and 3, representing Bathtub Gorge, Tooth Knob and Devil’s Peak respectively; B) Diagram showing generalised stratigraphic succession in the Flinders Ranges.
Material and methods
The studied material is reposited at the South Australian Museum in Adelaide. A total of 15 slabs were studied, most containing multiple specimens. Thirty-one specimens chosen based on quality of preservation were measured and included in the data processed for this report. These came from the Bathtub Gorge, Tooth Knob and Devil’s Peak localities of South Australia (1-3, respectively in Fig. 1A), from beds of the recently-proposed Nilpena Sandstone Member (Gehling et al., 2019), late Ediacaran Period (Fig. 1B). All fossil material is catalogued at the South Australian Museum Palaeontological Collections (specimen number prefix SAM P). Quantitative data was collected using digital callipers, protractor and a measuring string. Photography was done using low angle light with an Olympus D4 Tough digital camera and Canon EOS 50D SLR, with lenses EF-S 17-85 mm and EF 50 mm. Figures were produced with Adobe Photoshop CS3, Microsoft Excel and R (R Core Team, 2021).
Measured physical parameters were standardised, as illustrated by the simplified sketch in Figure 2, where we label the round end as the 'spoon’ and the sharp end as the ’knife’ and defined an arbitrary left and right bank when the fossil is viewed from the latter to the former. Unit length L was taken in millimetres from central axis to the end of the unit, unit width W was taken in millimetres from lateral groove to lateral groove, and unit angle 0 was measured as degrees from the next knife-ward axial junction (0 degrees for reference). Minimum specimen length was measured in millimetres with string, and minimum units were counted along the impression’s left bank and documented alongside. Both left and right banks were measured initially, as unit 1, 2, 3... and 1.5, 2.5, 3.5... respectively to represent the offset positions of the units, although for simplicity of data presentation, only left bank measurements were included in this report. An exception to this was for specimens measured from latex moulds, which were obviously inverted and so instead were measured along their right bank, serving as an inverted facsimile of the actual left bank. Due to the long, sometimes sinuous nature of P. hanseni fossils, along with fragmentary specimens and poor quality of preservation in some cases, only specimens showing clear units were measured for unit number and dimensions, and only complete specimens preserved in context were documented along their full profile. The spoon and knife of several specimens were measured for the first 25 units only, and only when preservation quality permitted. Measuring with such a close eye allowed a detailed view of some very small surface features, which were later investigated using putty. All the quantitative data was entered into Microsoft Excel and statistical analyses of the relationships between variables were conducted in R.
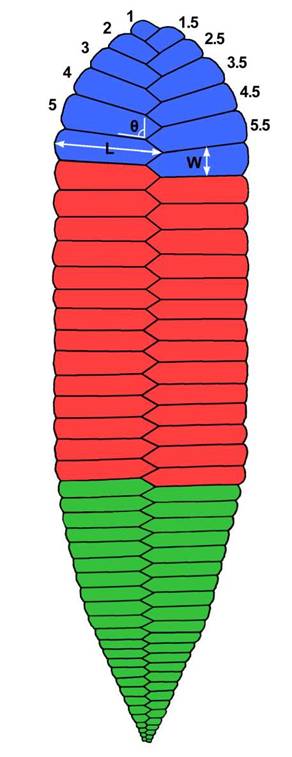
Figura 2: Diagrama esquemático de Phyllozpon hanseni indicando los parámetros biométricos investigados. Sistema de numeración de unidades desde el extremo. Código de colores: azul = región ‘cuchara’, rojo = región ‘tronco’ y verde = región ‘cuchillo’. / Figure 2. Schematic diagram of Phyllozoon hanseni outlining the biometric parameters investigated. Unit numbering system used from the terminus. Colour code: blue = ‘spoon’ region, red = ‘trunk’ region, and green = ‘knife’ region.
Systematic PalaeontologyPhylhzoon Jenkins and Gehling 1978 Type species: Phyllozoon hanseni Jenkins and Gehling 1978, by original designation and monotypy. Phylhzoon hanseni Jenkins and Gehling 1978 figs 5-8, S1, S3-S4.
1978 Phylhzoon hanseni Jenkins and Gehling, p. 357358, fig. 7.
1991 PhyUozoon hanseni Jenkins and Gehling; Ge-hling, pl. 3, fig. 2.
1992 Phylhzoon Jenkins and Gehling; Runnegar, fig. 3.10.
1994 Phylhzoon hanseni Jenkins and Gehling; Runne-gar, fig. 3.
2003 Phylozoon hanseni Jenkins and Gehling; Seila-cher, Grazhdankin and Legouta, p. 45-46, fig. 5. 2005 Phyllozoon Jenkins and Gehling; Gehling, Dro-ser, Jensen and Runnegar, p. 51, fig. 5.
2005 Phyllozoon Jenkins and Gehling; Seilacher, Bua-tois and Mangano, pp. 328-329, fig. 4.
2007 Phyllozoon Jenkins and Gehling; Seilacher, p, 178, pl. 62.
2007 Phyllozoon hanseni Jenkins and Gehling; Jenkins and Nedin, p. 209, fig. 8c.
2007 Phyllozoon hanseni Jenkins and Gehling; Reta-llack, p.17, fig. 7.
Material: SAM P19508A (holotype, Fig. 3A) and P19508B-C (paratypes), all on same slab, Nilpena Sandstone Member, Rawnsley Quartzite, Devil’s Peak, south of Quorn, southern Flinders Ranges (Jenkins and Gehling 1978, fig. 7). Additional 30 specimens in the SAM collections from Bathtub Gorge, Tooth Knob, Nilpena Ediacara National Park and Chace Range (pers. obs.).
Diagnosis: “Frond leaf-like, resembling Pteridinium simplex in its incised zig-zag median line and almost evenly-spaced, repetitive lateral grooves, but differing in that the organism was evidently more or less of planar or two dimensional form, not with three wings extending from the axis; incised axis and lateral grooves evidently representing a resistive skeleton which lay in foliate base of frond; lateral grooves extending form axis at about 70°, with their ends curved outwards; spacing of grooves tending to become less at (?) proximal end of frond.” (Jenkins and Gehling 1978, p. 357).
Description: Phyllozoon hanseni is an elongate, frond-like, soft-bodied fossil impression. Specimens range from just under 50 mm to more than 250 mm in length, and 20-40 mm wide, occasionally reaching up to 50 mm wide. The fossil generally consists of serially-arranged, hemicylindrical units that meet the axis at a regular sawtooth junction, displaying glide-reflection symmetry. P. hanseni has two termini that are morphologically distinct, with one made up of thicker units preserved at acute angles to the axis, transitioning gradually to the other end consisting of gradually thinner, shorter units arranged largely perpendicular to the axis.
Results
The holotype, South Australian Museum SAM P19508A (Fig. 3A), displays a complete specimen connected to what is here interpreted as an overlying partial specimen but could be mistaken for a case of branching, which is not observed in other specimens. Complete specimens where all the units are measurable are uncommon. The holotype was measured, along with another two complete and much smaller specimens, SAM P59027 (Fig. 3B) and SAM P59026 (Fig. 3C). From the measurement of these 3 full-length profiles (Fig. 3D), a consistent profile can be recognised, with unit length showing a steep increase at the spoon end, remaining similar for most of the trunk before showing a gradual decrease in unit length at about two thirds along the full profile to the knife.
Specimens SAM P35687-8 show what appears to be merging of units between the two adjacent individuals (arrows, Fig. 3E). On another slab, SAM P57687 is a significantly curved specimen that shows unit expansion and compression, with ‘lifting’ on the inside of the bend (arrow, Fig. 3F), in what appears to be a separation from the microbial mat in the seafloor, and thus lack of preservation. There is also one specimen that shows what could be interpreted as budding of globular chains at the edge of units along a consistent cleavage plane (SAM P59025, Fig. 3G) which has superficial resemblance to Funisia (Fig. 3H).
A histogram of specimen lengths was produced (Fig. 4A) for 17 individuals that were objectively measurable along full length and width of the specimen, preserved in context. Using a Shapiro-Wilk normality test (Shapiro and Wilk, 1965) of the specimen lengths produces a non-normal distribution (n = 17, M = 231 mm, W = 0.77, p = 0.0009), with a negative skew. Other analyses found a positive relationship (n = 17, R2 = 0.58, p < 0.0001) between the specimen length and unit number (Fig. 4B) when fitted to a robust generalised linear model with Poisson variance function. The plot in Figure 4C displays a positive linear relationship between specimen length and specimen width (n = 17, R2 = 0.39, p = 0.007) based on a reduced major axis (model IT) regression. Further characterisation of the spoon and knife was pursued, so unit length, width, and 0 angle (Fig. 2) of ten of each were measured for 25 units from the terminus and plotted graphically (Fig. 5). Generalised additive mixed model analysis, with individual specimen variation characterised as random effects, demonstrated monotonic but non-linear patterns across units for each length or angle measure in both the spoon and knife. The exception was for the angle measurement for the knife (Fig. 5F), which decreased linearly across units, despite appearing to hold a lower angle threshold of around 90° to the axis.
Discussion
There is considerable literature on the environmental setting of the South Australian Ediacara Biota in general (e.g, Gehling and Dro-ser, 2012; Reid et al. 2020), with some focused on the units where Phyllozoon has been collected (Gehling et al, 2019). The consensus around these units is that it was a shallow marine environment where, despite abundant energy, there was enough time for the microbial mat to grow and upon burial, via early permineralization, replicate the soft-bodied organisms on those seafloors and its sedimentary structures, like ripples, which should otherwise be scoured away by the very currents that formed them (Tarhan et al., 2017). Like with many other Ediacaran taxa, there is some debate as to what exactly P. hanseni was, although most seem to agree that it was closely related to Pteridinium Gurich, 1933 and Ernietta Pflug, 1966, members of the Nama Assemblage from Namibia, partially due to the similarities in quilt-like units, and despite the 3-dimensional preservation mode of the latter (Elliott et al., 2016). This would place Phyllo%oon within the Erniettomorpha (Pflug, 1972). Higher-level phylogenetic affiliation for Phyllo%oon, as with a number of Ediacara Biota members, remains undetermined, and falls outside the scope of this paper. However, an interesting feature recognised here for the first time, which could be of some phylogenetic value, is that the body units in P. hanseni always start on the same side: left of the hyporelief fossil (right side in life), as we look from knife to spoon (Fig. 2), implying that Phyllo%oon started from the spoon end and added units on the knife end.
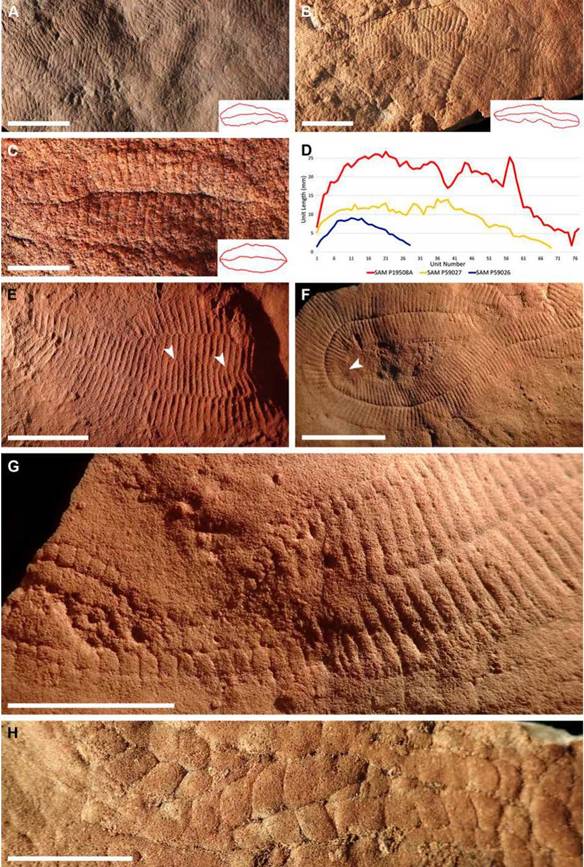
Figura 3: Phyllozoon hanseni Jenkins y Gehling 1978, del Ediacárico de la Sierra de Flinders, Australia del Sur. A) Vista completa del holotipo SAM P19508A (Escala: 50 mm); B) Vista completa de SAM P59027 y P59028 (Escala: 25 mm); C) Vista completa de SAM P59026 (Escala: 10 mm); D) Diagrama mostrando la variación de longitud de las unidades, a lo largo del perfil de los tres ejemplares de P. hanseni en Figs 3A, B y C; E) Molde de látex de SAM P35687 y SAM P 35688, mostrando las unidades fusionadas (flechas) entre estos dos ejemplares (Escala: 25 mm); F) Ejemplar SAM P57687, con unidades difusas (flecha, elevadas sobre el tapiz microbiano) en el borde interior (Escala: 50 mm); G) Gemas globulares concatenadas, formadas en el extremo distal de las unidades corporales del ejemplar SAM P59026 (Escala: 25 mm); H) Unidades globulares concatenadas de Funisia SAM P42371, para comparación con Fig. 3G (Escala: 25 mm) / Figure 3. Phyllozoon hanseniJenkins and Gehling 1978, from the Ediacaran of the Flinders Ranges, South Australia. A) Full length photograph of holotype SAM P19508A (Scale Bar: 50 mm); B) Full length photograph of SAM P59027 and P59028 (Scale Bar: 25 mm); C) Full length photograph of SAM P59026 (Scale Bar: 10 mm); D) Diagram illustrating unit length changes along the profile of the three specimens of P. hanseni from Figures 3A, 3B, and 3C; E) Latex mould of SAM P35687 and SAM P 35688, displaying merged units (arrows) between these two specimens (Scale Bar: 25 mm); F) Large, curved specimen SAM P57687, with diffuse units (arrow, lifted above the microbial mat) in the inside edge of the bend (Scale Bar: 50 mm); G) Chained budding features forming along the distal end of body units on specimen SAM P59026 (Scale Bar: 25 mm); H) Globular chained segments of Funisia SAM P42371, for comparison with Fig. 3G (Scale Bar: 25 mm).
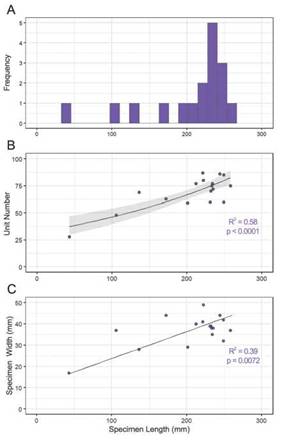
Figura 4: A) Histograma que muestra la distribución no-normalizada y de sesgo negativo (M = 231 mm, W = 0.77, p = 0.0009) de los 17 ejemplares completos medidos de P. hanseni; B) Gráfico de dispersión que muestra una correlación positiva entre la longitud del ejemplar y el número de unidades basado en modelo lineal generalizado robusto (n = 17, intervalo de confianza representado en gris); C) Gráfico de dispersión que muestra longitud frente a anchura de los ejemplares (n = 17) basado en una regresión de modelo II (errores en ambas variables). / Figure 4. A) Histogram displaying the non-normal, negative-skewed size distribution (M = 231 mm, W = 0.77, p = 0.0009) of the 17 P. hanseni specimens measured; B) Scatter plot showing a positive relationship between specimen length and unit number based on a robust generalised linear model (n = 17, 95% confidence interval represented in grey); C) Scatter plot mapping specimen length against specimen width (n = 17) based on a model II regression (errors in both variables).
There are some novel features of interest (Fig. 3) which may present several appropriate directions for future research. Firstly, the junction on SAM P19508A (Fig. 3A) should be investigated in detail, as although interpreted as a partial specimen overlap, further analysis should allow to totally refute a case of branching. Additionally, a study of new material as it becomes available could provide insight as to whether the merged units between specimens SAM P35687 and SAM P35688 (Fig 3E) is an element of chance or a biological feature, where fusion of units from independent specimens occurs. The chains trailing from the edges of Phyllo%oon on SAM P59025 (Fig. 3G) should also be further analysed and compared with specimens of the resembling Funisia (Fig. 3H). This taxon varies widely in size, and as mentioned earlier, shares the same positive hyporelief preservation, while also being known from wave-base and sheet-flow sands facies, in which Phyllo%oon and Aulo-zpon’ also occur (Reid et al., 2020).
A negatively skewed, non-normal distribution was produced by the specimen length data (Fig. 4A), although some of this may be a result of preservation or collection bias, as larger specimens may have a better preservation potential. Figure 4B seems to suggest that P. hanseni grows by increasing both unit number and size which could prove useful as a predictor in similar future analyses of many more partial specimens and could be further developed upon recording more full-length specimens of different size ranges unearthed through future excavations. Figure 4C corroborates observations that it also grows only a fraction wider through its lifetime, but substantially longer. This information has the potential to yield further insights about mode of life. The spoon and knife generalised additive mixed model analyses (Fig. 5) illustrate a stark contrast between the spoon’s rounded form and the knife’s linearity, which could speak of physical or structural constraints within the body plan of these organisms. Perhaps some level of turgor or rigidity held smaller knife-ward units straight out perpendicular to the longitudinal axis. These trends could also hold functional implications: as P. hanseni was a benthic organism, the elongated teardrop shape - the rounded spoon -, steadily tapering toward the knife could have reduced turbulence and pressure drag, so that currents had less chance of damaging delicate structures, while the bulbous spoon end could provide a larger surface area to serve as anchor.
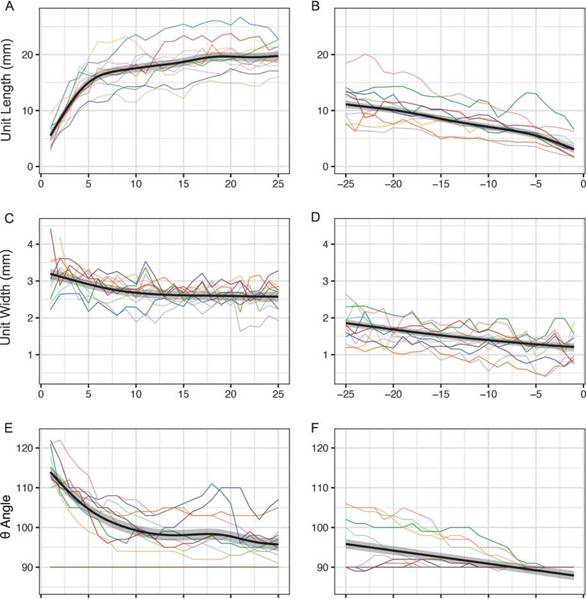
Figura 5: Gráficos del modelo mixto acumulativo generalizado de: A) Longitud de las primeras 25 unidades de la ‘cuchara’ de 10 ejemplares; B) Longitud de las últimas 25 unidades del ‘cuchillo’ de 10 ejemplares; C) Ancho de las primeras 25 unidades de la ‘cuchara’ de 10 ejemplares; D) Ancho de las últimas 25 unidades del ‘cuchillo’ de 10 ejemplares; E) Ángulo de las primeras 25 unidades de la ‘cuchara’ de 10 ejemplares; F) Ángulo de las últimas 25 unidades del ‘cuchillo’ de 10 ejemplares; línea marrón en E y F indica 90° respecto al eje sagital de P. hanseni. / Figure 5. Generalised additive mixed model plot of: A) Length of the first 25 ‘spoon’ units of 10 specimens; B) Length of the last 25 ‘knife’ units of 10 specimens; C) Width of the first 25 ‘spoon’ units of 10 specimens; D) Width of the last 25 ‘knife’ units of 10 specimens; E) Angle of the first 25 ‘spoon’ units of 10 specimens; F) Angle of the last 25 ‘knife’ units of 10 specimens; brown line in E and F indicates 90° to the sagittal axis of P. hanseni.
Conclusion
This preliminary study on Phyllozoon hanseni has provided new information on this relatively rare Ediacaran taxon, only known from Australia. From a literature review, first-hand analysis of the fossils, to the features and measurements photographed and documented, this investigation posits that P. hanseni was possibly a benthic organism, despite lacking a singular central holdfast. It may have been made of a soft tubule matrix and may have reproduced by budding from these units/tubules. With the superficial similarities between these budding units with Funisia, their shared positive hyporelief preservation and shared presence in the wave-base and sheet flow sands facies, there may be some relationship between the two taxa yet to be realised. Phyllopoon grew from the oldest spoon region to the youngest knife region, adding units consecutively at the knife tip. Some individual specimens recorded more than 50 mm wide and longer than 300 mm, and as they grew in total length, they grew in unit number, and as units grew in length, they also grew slightly thicker. Most interestingly, the organism’s width appears to be far more tightly constrained than its body length, with specimen widths appearing to grow towards a threshold. New material is being unearthed every year, particularly from the Nilpena site and will undoubtedly continue to provide further insights into the morphology of Phyllo%oon hanseni.
Acknowledgements
We would like to thank South Australian Museum’s Prof. Jim Gehling and SAM Palaeontological Collection Manager Dr. Mary-Anne Binnie for providing the opportunity to access and study the specimens on display and in the collections, as well as for their first-hand accounts, guidance, and dedication to the project. AMT would also like to acknowledge Miss Tory Botha for her statistical support, coding assistance, and sharing of helpful resources, as well as Dr. James Holmes, for lengthy scientific discussions. Thanks to Dr. Susana Heredia (CONICET, Universidad Nacional de San Juan, Argentina) for her careful revision of the manuscript. Additional gratitude is owed to Mrs. Jess Turner who is always very supportive and happy to converse about deep time and Earth history, and also to Mr. Ray Barnes for his diligent assistance with data entry. Lastly, we wish to extend a special thanks to Sandy’s Memorial Trust for their financial contribution to AMT’s summer research project through the South Australian Museum.

