










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgriscientia
versión On-line ISSN 1668-298X
Agriscientia vol.32 no.1 Córdoba jun. 2015
ARTÍCULOS
Dinámica de la comunidad de artrópodos asociada a cultivos de frutilla con plantas de diferentes edades en el noroeste de la Argentina
Olivo, V.I.; J.A. Corronca y A.X. González Reyes
V.I. Olivo: Cátedra de Zoología Agrícola, Facultad de Ciencias Naturales, Universidad Nacional de Salta - IEBI, Av. Bolivia 5150, Salta, Argentina. J.A. Corronca: CONICET - Cátedra Artrópodos, Facultad de Ciencias Naturales, Universidad Nacional de Salta - IEBI, Av. Bolivia 5150, Salta, Argentina. A.X. González Reyes: Cátedra de Invertebrados, Facultad de Ciencias Naturales, Universidad Nacional de Salta - IEBI, Av. Bolivia 5150, Salta, Argentina. Correspondencia a: veroolivo@iebi.com.ar
RESUMEN
Dentro de un manejo de monocultivo de frutilla, el diseño de parcelas con plantas de distintos años de producción, lleva al sistema a tener mayor estabilidad y resiliencia y afecta de distinta manera tanto a las plagas como a sus enemigos naturales. El objetivo del trabajo fue evaluar si existen diferencias en la diversidad y en la estructura de la comunidad de artrópodos, considerando su variación temporal de acuerdo con la fenología del cultivo de frutilla en el período de floración-fructificación, en parcelas de similares dimensiones con plantas de iguales y distintas edades. La comunidad de artrópodos fue diferente entre años analizados, siendo más diversa la comunidad de artrópodos cuando el cultivo de frutilla presentaba plantas de diferentes edades en el mismo minifundio. Las plagas principales y los enemigos naturales en este estudio variaron entre años y entre las parcelas de plantas de distintas edades. Se concluye que la comunidad de artrópodos en un cultivo de frutilla en un mismo predio, varía con los años y con la fenología del cultivo y entre parcelas con plantas con edades diferentes de implantación.
Palabras clave: Plagas; Enemigos naturales; Frutilla; Fenología; Minifundio.
Dynamic of the arthropods community associated to strawberry crops with different plant ages in the North-West of Argentine.
SUMMARY
In a strawberry monoculture management, the design of the plots with plants of different years of production, would lead to greater stability and resilience in the system, affecting differently both pests and their natural enemies. The objective of this work was to assess whether there are differences in the diversity and structure of the arthropod community, considering its temporal variation according to strawberry crop phenology in the fowering and fruiting periods in plots of similar dimensions with plants of the same and of different ages. The arthropod community was different in composition among the years analyzed. The most diverse community of arthropods was observed when the crop had strawberry plants of different ages in the same smallholding. The main pests and natural enemies present in this study varied among years and plots of plants of different ages. The results lead us to suggest that arthropod community in a crop of strawberries in the same field varies with age and plant crop phenology and among plots with different ages of implantation.
Key words: Pests; Natural enemies; Strawberry; Phenology; Smallholding.
Fecha de recepción: 19/06/14;
fecha de aceptación: 08/06/15
INTRODUCCIÓN
Todos los agroecosistemas son dinámicos y están sujetos a diferentes tipos de manejo, de manera que los arreglos de cultivos en el tiempo y en el espacio cambian continuamente de acuerdo con factores biológicos, socioeconómicos y ambientales. Tales variaciones determinan el grado de heterogeneidad característica de cada paisaje agrícola de una región, la que a su vez condiciona el tipo de biodiversidad presente, que a su vez puede o no beneficiar la protección de cultivos en agroecosistemas particulares. Los artrópodos son un componente dominante de la cadena trófica de los herbívoros, carnívoros y detritívoros en la mayoría de los ecosistemas terrestres. Varias hipótesis sostienen que su abundancia se ve influenciada por la diversidad de las plantas (Perner et al., 2005). Sin embargo, autores como Haddad et al. (2001) resaltaron en sus estudios que aquella está influenciada más por los efectos de las características funcionales, estructurales y fenológicas de las plantas que por el número de especies cultivadas.
Si bien los monocultivos son ambientes poco favorables para los enemigos naturales de las plagas, debido a los altos niveles de perturbación y a la falta de infraestructura ecológica, el mayor desafío al manejar un sistema sucesional consiste en aprender a introducir perturbaciones de forma que estimulen la productividad del sistema, por un lado, y provean resistencia al cambio y la variación, por el otro. Esto se puede lograr de varias maneras, de acuerdo con las condiciones ambientales locales, la estructura de los ecosistemas naturales maduros normalmente presentes y la posibilidad de mantener las modificaciones en el largo plazo (Altieri & Nicholls, 2004). La resiliencia de un sistema es medida por la magnitud de perturbaciones que pueden ser absorbidas por el sistema antes de que sea reorganizado con diferentes variables y procesos (Holling, 1973). La heterogeneidad de un sistema está dada por el cambio de estructura de las plantas y será mayor cuando, en un segundo año, consiga mayor estabilidad debido a la variación física de aquellas. Es así que, dentro de un manejo de monocultivo, el diseño de parcelas con plantas de distintos años de producción (que le confiere heterogeneidad interna), llevará al sistema a tener mayor estabilidad y resiliencia, como así también afectará de distinta manera las relaciones tróficas entre los distintos niveles.
Las explotaciones agrícolas minifundistas (hasta 5 hectáreas) constituyen la mitad de los campos en la agricultura latinoamericana (Altieri, 1999), y el modelo se basa en cultivos agrícolas intensivos que generan un importante impacto sobre el ambiente. Son espacios adecuados para realizar estudios agroecológicos, dejando de lado los experimentos controlados que muchas veces no reflejan la realidad del campo.
El cultivo de frutilla es un cultivo de importancia social y económica, pues genera gran cantidad de puestos de trabajo -la mayoría de los productores son pequeños y familiares-, produce divisas a través de las exportaciones y se puede industrializar, logrando productos con mayor valor agregado (Proyecto INTA cartera 2009-2012). En la Argentina se producen 13.000 toneladas anuales de frutillas en un área de 1000 ha, de las cuales 40% corresponde a la provincia de Santa Fe, 30% a la provincia de Tucumán y el resto a otras zonas productoras. En el caso de las provincias de Salta y Jujuy, es de 300 tn, con un rendimiento de 10 tn/ha y una superficie sembrada de 30 ha (Senasa, 2009).
Aunque la planta de frutilla es una herbácea perenne, se produce como una planta bianual ya que la calidad y el rendimiento son máximos en el primero y segundo año. La mayoría de las plagas de la frutilla son polífagas y sus enemigos naturales comúnmente están presentes en el cultivo y en otras plantas hospedadoras cercanas. El manejo del cultivo de frutilla, que incluye métodos de protección y otros, como la esterilización del suelo y las aplicaciones de plaguicidas, tiene influencia profunda sobre las plagas y el complejo de enemigos naturales; sin embargo, sus efectos no han sido aún cuantificados de manera adecuada (Cross et al., 2001).
El objetivo de este trabajo fue evaluar si existen diferencias en la diversidad, y en la estructura de la comunidad de artrópodos, considerando su variación temporal de acuerdo con la fenología del cultivo de frutilla en el período de floración-fructificación, en parcelas de similares dimensiones con plantas de las mismas y diferentes edades.
MATERIALES Y MÉTODOS
Área de estudio
El estudio se llevó a cabo durante 2002 y 2004 en una finca minifundista de 4,5 ha de superficie, en la localidad de Vaqueros (24°42´S, 65°24´W), Salta, Argentina. La finca estudiada presentó el 75% del área cultivable con parcelas de frutilla y el resto con parcelas con distinta rotación de cultivos hasta barbecho. Todo el minifundio estuvo rodeado por una importante barrera rompevientos de vegetación nativa del Chaco Serrano salteño. Durante el año 2002, la parcela de frutilla (en adelante FR-2002) (aproximadamente 3 ha) mantuvo plantas de la misma edad y estuvo rodeada por una parcela de cebolla y de arveja, totalizando casi 1 ha cultivada. En 2004, el cultivo de frutilla (FR-2204) (3 ha) estuvo representado por dos parcelas de similar tamaño pero con plantas de diferente edades, una recién implantadas (en adelante Fr2004P1) y otra de plantas de segundo año de producción (en adelante Fr2004P2), separadas por 2 metros sin cultivar. Acompañando estas parcelas con frutilla, durante el período de estudio hubo una parcela de no más de 0,4 ha con arveja, y el resto del minifundio (0,6 ha) permaneció en barbecho. La parcela de frutilla Fr2004P2 recibió una sola aplicación de agroquímicos y presentó malezas en los últimos momentos del muestreo. En los dos años de producción la plantación de frutilla se realizó en camellones estrechos con plásticos negros, con dos filas de plantas en cada uno de ellos con una línea de riego por goteo.
Muestreo
Los muestreos se realizaron desde julio hasta noviembre de 2002 y 2004, período de floración-fructificación de la frutilla. Estos fueron quincenales y se realizaron con un aspirador manual G-Vac (garden vaccum), para los artrópodos de la parte aérea de las plantas, y con trampas de caída (pit-fall) para la fauna epigea; se consideraron 15 muestras al azar con cada metodología en las parcelas cultivadas. En cada punto de muestreo se colocó una trampa de caída de 10,5 cm (Ø), con líquido conservante (etilenglicol: agua en solución 1:10, con gotas de detergente) y se tomó una muestra de fauna sobre la vegetación con un G-Vac; cada muestra consistió en la succión de ocho plantas completas de frutilla por el término de 1 minuto. El material obtenido por cada método fue separado e identificado en una primera etapa a nivel de morfoespecie, apoyado por una base de imágenes digitales generadas por el software Taxis 3.5 (Meyke, 2003). Posteriormente el material fue separado en órdenes y familias y clasificado en grupos funcionales. Las categorías tróficas de las distintas especies fueron asignadas con base en el conocimiento previo de su biología y, en algunos casos, considerando el o los grupos tróficos que dominan la familia a la que pertenecen. Cuando fue posible, se identificó el material a nivel específico por medio de claves taxonómicas disponibles y la comparación con material de colecciones de referencia de la región depositadas en la Cátedra de Zoología Agrícola de la Facultad de Ciencias Naturales, Universidad Nacional de Salta.
La fauna de artrópodos se separó en tres gremios: fitófagos, depredadores y parasitoides. Se generaron gráficas de su abundancia a lo largo del tiempo, relacionándolas con los momentos de aplicaciones de agroquímicos en cada parcela estudiada. El productor realizó las siguientes aplicaciones de agroquímicos durante el período en estudio: durante el año 2002, tres aplicaciones, dos con Cipermectrina (insecticida piretroide de amplio espectro) y uno con Vertimec (insecticida y acaricida de origen natural); en 2004, en la parcela Fr2004P1, cuatro aplicaciones, una con Avermectina, Cipermectrina-azufre, azufre y Vertimec, y en Fr2004P2, una única aplicación de Cipermectrina-azufre.
Análisis de datos
Para conocer y comparar la artropodofauna asociada y su variación en el cultivo de frutilla, se generaron gráficas de la variación total de la abundancia de artrópodos en los sitios y a lo largo de período estudiado. Se generaron curvas de acumulación de especies basadas en muestras para cada sitio usando el programa EstimateS 7.0 (Colwell, 2004). Las comparaciones de la diversidad de las comunidades de artrópodos se hicieron teniendo en cuenta medidas sencillas y comunes de la diversidad, como el índice de riqueza de especies (S), abundancia relativa (n), índice de diversidad Shannon (H '), el índice de dominancia de Simpson & Berger-Parker (D) y la equidad (J) (Krebs, 1999; Magurran, 2004). Se empleó el método de bootstrap (Krebs, 1999) para comparar los valores obtenidos para los parámetros estudiados. Estos análisis se realizaron utilizando el software PAST 1.18 (Hammer et al., 2003).
En el análisis de la fauna de artrópodos con respecto a los estados fenológicos de la frutilla en las parcelas estudiadas, se utilizó un escalamiento no métrico multidimensional (NMS) a través del programa PC-ORD (McCune & Grace, 2002). Para ello se realizó una matriz primaria con los datos de abundancia de las especies de artrópodos de acuerdo a la fenología del cultivo y se siguieron los procedimientos sugeridos por McCune & Grace (2002). En la primera corrida, se usó el modo Autopilot slow and thorough (programa PC-ORD ver.6.0), con Sorensen como medida de distancia, 500 iteraciones, configuración de comienzo al azar, 250 corridas con datos reales y 1 en la reducción de la dimensionalidad para cada ciclo. Para evaluar cuando el NMS extrajo ejes más robustos que lo esperado por azar, se aplicó el test de Montecarlo. Este análisis se realizó reiteradamente (sin opción Autopilot) para confirmar el resultado antes mencionado, con la intención de disminuir el estrés al mínimo valor posible, pero en todos los casos el valor obtenido se mantuvo invariable. Se realizó un análisis de permutación de múltiple respuesta (MRPP), utilizando el índice de Sorensen (=Bray-Curtis) como medida de similitud, a fin de evaluar si la comunidad mostraba cambios en su composición en los años estudiados y durante los distintas etapas fenológicas del cultivo en cada año, para ello, se usó el programa PC-ORD ver.6.0 (McCune & Grace, 2002).
RESULTADOS
Diversidad de artrópodos
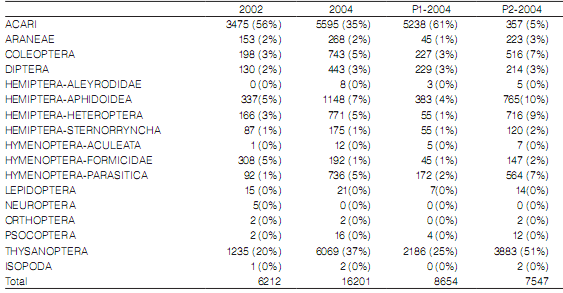
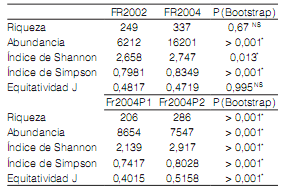
Se recolectaron 22.413 artrópodos correspondientes a 78 familias en el año 2002 y 94 en 2004 (Fr2002: S=249 y n=6212; Fr2004: S=337, n=16.201). Se observó una diferencia en la composición de los órdenes de artrópodos y sus abundancias en las parcelas estudiadas (Tabla 1). Acari y Thysanoptera fueron los órdenes de artrópodos más abundantes durante el estudio y ambos mostraron abundancias similares en 2004, pero no en 2002. Los artrópodos recolectados durante 2004 se distribuyeron de la siguiente manera: 8654 individuos correspondientes a Fr2004-P1 (S=206 de 76 familias) y 7547 a Fr2004-P2 (S=286 de 85 familias). Los ácaros presentaron baja abundancia en P2-2004, no así en P1-2004, donde fueron dominantes.
Tabla 1. Abundancias absolutas y porcentajes relativos de grupos de artrópodos, comparados entre años y entre parcelas de plantas de distintas edades.
Según las curvas de acumulación de especies por muestras, existen diferencias estadísticamente significativas en la riqueza de especies entre los años estudiados (Figura 1A), siendo más diversa la fauna del 2004. Los valores de los índices de diversidad de Shannon y de dominancia de Simpson muestran diferencias estadísticamente significativas (p<0,05) entre años; el cultivo de frutilla de 2002 es de menor dominancia. Al considerar en 2004 las parcelas por separado, el cultivo de segundo año de producción presentó mayor diversidad en cuanto a riqueza de especies (Figura 1B y Tabla 2).
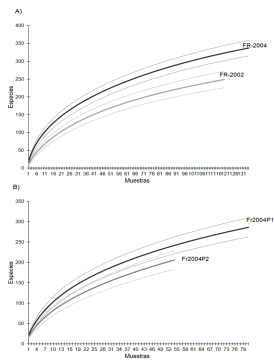
Figura 1. Curvas de acumulación de especies por muestra. A. 2002 vs 2004, B. P1-2004 vs P2-2004. Líneas punteadas indican el intervalo de confianza.
Tabla 2. Valores de riqueza, abundancias e índices de diversidad en las parcelas estudiadas. * p<0,05, NS p>0,05.
Las comunidades de artrópodos fueron distintas en los dos años analizados, según lo muestra el análisis de NMS (Figura 2) y con diferencias estadísticamente significativas (MRPP: A= 0,0508, p= 0,038). El NMS presenta el mínimo estrés total de 8,23, por lo que existe una buena ordenación sin riesgo real de falsa inferencia y, por lo tanto, se considera satisfactorio. El test de Montecarlo muestra que existen diferencias altamente significativas (p≤0,04) entre los valores de estrés observado para los ejes extraídos por el NMS. La configuración final revela que las comunidades de artrópodos fueron distintas en los dos años.
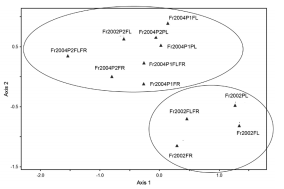
Figura 2. Ordenamiento por medio de un análisis de escalamiento multidimensional no métrico (NMD) de las etapas fenológicas del cultivo de frutilla. Fr2002Pl: frutilla 2002 en estado vegetativo; Fr2002Fl: frutilla 2002 en floración; Fr2002FLFR: frutilla 2002 en floración/fructificación; Fr2002FR: frutilla 2002 en fructificación; Fr2004P1Pl: frutilla 2004 parcela 1 en estado vegetativo; Fr2004P1Fl: frutilla 2004 parcela 1 en floración; Fr2004P1FLFR: frutilla 2004 parcela 1 en floración/fructificación; Fr2004P1FR: frutilla 2004 parcela 1 en fructificación; Fr2004P1FR: frutilla 2004 parcela 1 en fructificación; Fr2004P2Pl: frutilla 2004 parcela 2 en estado vegetativo; Fr2004P2Fl: frutilla 2004 parcela 2 en floración; Fr2004P2FLFR: frutilla 2004 parcela 2 en floración/fructificación; Fr2004P2FR: frutilla 2004 parcela 2 en fructificación
Al comparar entre sí, a través de MRPP, las dos parcelas de 2004, se observa que también existen diferencias significativas entre las comunidades de artrópodos arriba mencionadas (A=0,035, p=0,0254).
La abundancia de artrópodos en las parcelas cultivadas en los dos años registró un aumento marcado en el inicio del período fructificación (Figura 3, muestreos de octubre) y en todos los casos los valores del análisis de MRPP entre floración y fructificación mostraron diferencias estadísticamente significativas (Fr2004-P2: A=0,1262, p=0,007; Fr2004-P1: A=0,2476, p=0,0244; Fr2002: A= 0,1228, p=0,01058).
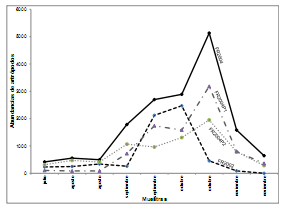
Figura 3. Abundancia temporal de artrópodos en los dos años analizados y en las dos parcelas del 2004.
El patrón de variación de la abundancia de artrópodos al comparar parcelas del año 2004 fue casi similar, con mayor abundancia relativa de artrópodos en Fr2004-P2, debido a un incremento considerable de Caliothrips phaseoli Hood (Figura 3). La diferencia en el patrón de abundancia observado en los muestreos de septiembre se debe a que en Fr2004-P1 las plantas estaban en estado de crecimiento vegetativo y hubo un aumento considerable de ácaros.
Análisis de grupos funcionales
En ambos años muestreados los fitófagos mostraron un porcentaje similar, 85% del total de artrópodos recolectados, mientras que los parasitoides fueron los que más variaron en abundancia entre años, incrementando un 4% en 2004 con respecto a 2002. La variación a lo largo del tiempo de los grupos funcionales no fue similar entre años, especialmente en el caso de los parasitoides, que mostraron cambios marcados en 2004, siendo más abundantes que en 2002 (Figura 4A y B). En el caso de los fitófagos, salvo un aumento de la abundancia en el inicio de la floración, su variación fue similar: Caliothrips phaseoli (Hood), Tetranychusurticae (Koch), Aphis gossypii Glover, Myzus persicae (Sulzer, 1776) y Nysius sp. fueron las principales plagas registradas en el cultivo de frutilla en el Valle de Lerma, y mostraron cambios en su abundancias en los años estudiados. En 2002, T. urticae (Acari) y C. phaseoli (Thysanoptera) fueron dominantes (28% y 11%, respectivamente); en cambio, en 2004 lo fueron C. phaseoli (20%) y T. urticae (18%).
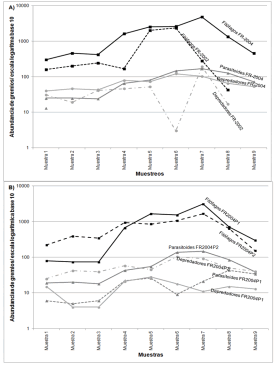
Figura 4. Variación temporal de grupos funcionales: A. 2002 vs 2004; B. FrP1-2004 vs FrP2-2004
Si se analiza el comportamiento de las principales plagas en cuanto a abundancia entre años, se pudo observar que no hubo un patrón definido: los ácaros aumentaron hacia el final del período floración-fructificación, los pulgones fueron abundantes en la etapa de crecimiento vegetativo o antes de la floración y las chinches fitófagas (Hemiptera Heteroptera) aumentaron gradualmente su abundancia con el tiempo (Figura 5 A). C. phaseoli y T. urticae mostraron un incremento desde la floración-fructificación y los pulgones una disminución en su abundancia (Fig. 5A y B).
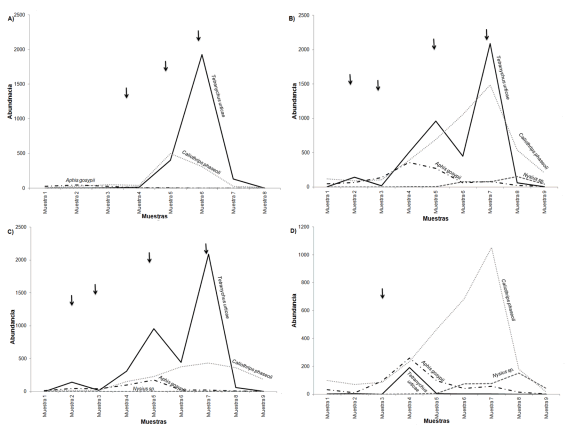
Figura 5. Variación temporal de principales plagas registradas en: A. Fr2002 B. Fr2004 C. Fr2004P1 D. Fr2004P2. Las flechas indican las prácticas de manejo realizadas por el productor.
En las parcelas con frutilla de diferentes edades se registraron las mismas plagas, pero sus fuctuaciones en abundancias fueron diferentes. Así en frutilla de primer año, T. urticae fue dominante mientras que C. phaseoli lo fue en las del segundo año de producción. A lo largo del tiempo, las especies de trips plagas y los pulgones (Hemiptera Fam. Aphididae) mostraron patrones casi similares de fluctuación temporal en ambas parcelas, no así los ácaros ni las chinches fitófagas (Figura 5C y D). Al comparar las parcelas con diferentes edades de implantación se observó que la composición del complejo de plagas fue diferente entre ellas.
Con respecto a los depredadores, durante el estudio se mantuvieron en niveles bajos de abundancia. Los principales depredadores registrados en 2002 fueron Orius insidiosus Say (Hemiptera Heteroptera Fam. Anthocoridae), Eriopis connexa (Germar) (Coleoptera Fam. Coccinellidae) y una especie de araña licósida; mientras que en el año 2004 fueron varias especies de arañas de la Familia Araneidae, coleópteros estafilínidos y una especie de chinche del género Geocoris (Hemiptera Heteroptera Fam. Geocoridae) (Figura 6A y B). Las chinches depredadoras mostraron un patrón similar en su abundancia entre años, mientras que no ocurrió lo mismo con las arañas. La composición de los depredadores y sus fluctuaciones fueron diferentes entre las parcelas con distintas edades, y mostraron ser los ácaros fitoseidos muy escasos (Figura 6C y D).
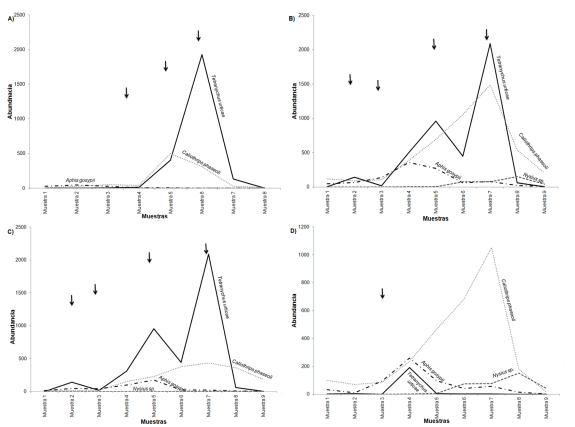
Figura 6. Variación temporal de principales depredadores registrados en A. Fr2002 B. Fr2004 C. FrP1-2004 y D. FrP2-2004. Las flechas indican las prácticas de manejo realizadas por el productor.
DISCUSIÓN
Al evaluar la diversidad sobre la base de la riqueza de especies (curvas de acumulación de especies), de la equidad y de la dominancia (índice de Simpson & Berger-Parker), y a pesar de que las parcelas recibieron aplicaciones de agroquímicos, se observa una gran diversidad de artrópodos en los dos años muestreados, al igual que lo encontrado por Lemme et al. (1996) en cultivos de frutilla de la provincia de Tucumán.
Además, la comunidad de artrópodos fue diferente en cuanto a composición en los años analizados, más diversa cuando el cultivo de frutilla presentó plantas de diferentes edades en el mismo minifundio. Estos resultados permiten inferir que la mayor heterogeneidad interna al contar con plantas de diferentes edades, implicó diferencias físicas estructurales en ambas parcelas, que puede explicar este incremento en la diversidad y abundancia de artrópodos. Además, esta idea está apoyada por los resultados obtenidos donde las parcelas de frutilla con plantas de segundo año mostraron una mayor diversidad, riqueza de especies y una comunidad más equitativa, en contraposición a la alta dominancia de algunas especies observadas en la parcela con plantas recién implantadas. La tendencia a la estabilización de la comunidad de artrópodos, sumada a las invasiones sucesivas de los hábitats adyacentes (Van Emden, 1965; Huffaker & Messenger, 1976; Altieri, 1994; Fry, 1995), posiblemente pueda explicar lo observado en la parcela de segundo año.
Al igual que la mayoría de los trabajos realizados en cultivos de frutilla (Sorensen et al., 1997; Autio et al., 2004; Greco et al., 2005; Rondon et al., 2005; Zalom et al., 2005), la arañuela roja Tetranychus urticae fue la plaga principal en el cultivo estudiado. En ambos años de estudio se registró la plaga, comportándose como clave en Fr2004-P1, no así en Fr2004P2 (aun cuando fue abundante). Ello puede deberse a que las hojas son menos apetecibles cuando son más viejas (Gomez-Moya & Ferragut, 2009) y los ácaros migran buscando plantas en mejor estado para depositar sus huevos. Por otro lado, los efectos de los plaguicidas disminuyen significativamente no sólo su enemigo natural por excelencia, ácaros fitosiedos (registrados en muy bajo número), como lo menciona Greco et al. (2004 y 2007), sino también a la gran mayoría de los enemigos naturales. La alta abundancia de T. urticae en la parcela con frutilla de primer año durante 2004 también puede explicarse por la rápida capacidad que tiene para desarrollar resistencia a plaguicidas en pocas generaciones (Stumpf et al., 2001; Stumpf & Nauen, 2002).
Varias especies de trips infestan los cultivos de frutilla (Coll et al., 2006; Ripa et al., 2009), y a diferencia de otros estudios, aquí la especie dominante fue C. phaseoli. Esto en contraposición a los estudios que encontraron a Frankliniella occidentalis (Zalom et al., 2005; Pinent et al., 2011); F. bispinosa (Rondon et al., 2005) y F. schultzei (Lefebvre et al., 2013), en igual estado fenológico del cultivo (floración-fructificación). Otras especies de trips también se registran en este estudio y en otros, lo que muestra que existe un complejo de trips que infestan las flores de frutilla (Autio et al., 2004); la especie que encuentre su óptimo en esas condiciones será la que prevalecerá sobre las otras del complejo, como lo fue C. phaseoli (especialmente en las plantas recién implantadas). Las abundancias obtenidas de las especies de trips en la parcela de segundo año, estuvieron influenciadas tanto por la baja aplicación de plaguicidas (sólo una de insecticida piretroide de amplio espectro) como por la aparición de malezas y la estabilidad del cultivo lograda por la no remoción de la parcela. Esto indica que las frecuencia de las especies plagas de trips está influenciada tanto por la presencia de otros cultivos y vegetación natural, como por el hecho de que los trips son fácilmente acarreados por el viento (Pinent et al., 2011).
El porcentaje general de enemigos naturales en el minifundio es marcadamente inferior al de los fitófagos, manteniéndose con valores similares entre años. La parcela con plantas de segundo año, 2004, presenta una mayor abundancia de enemigos naturales y esto se debe a que la comunidad de enemigos naturales tiende a establecerse y estabilizarse en el tiempo, como ocurre en los cultivos perennes, y también a una menor aplicación de plaguicidas.
Orius insidiosus, y en menor medida E. conexa y una especie de araña licósida, son los depredadores con mayor abundancia en las parcelas de 2002. Su incremento en abundancia responde al incremento de las plagas que controlan (Aphis gossypi y T. urticae) (Rondon et al., 2005). El complejo de enemigos naturales dominantes en el 2004 es diferente, sumándose a O. insidiosus, una especie de Geocoris y una araña de la familia Araneidae y algunos coléopteros estafilínidos en menor número. Los resultados muestran que el complejo de enemigos naturales en las parcelas con plantas de frutilla de diferentes edades varía en la composición de especies. Así Geocoris sp. acompaña el incremento de su presa (T. urticae) en la parcelas con plantas de primer año, mientras que O. insidiosus lo hace siguiendo a C. phaseoli. Estos resultados no concuerdan con estudios en los que las chinches depredadoras prefieren a losácaros más que a otros fitófagos (Rondon et al., 2004) y lo reportado por Wright (2001), Richards & Schmidt (1996) y Laing (1969), en que Orius acompañó la densidad poblacional de T. urticae. En cambio, los resultados del presente trabajo coinciden con los obtenidos por Saini et al. (2003) en que O. insidiosus es la especie más frecuente y abundante que controla trips. En este sentido Lefebvre et al. (2013) han iniciado estudios tendientes a implementar su utilización como agente de biocontrol.
La comunidad de arañas, representada en este estudio, es poco abundante debido a su alta susceptibilidad a la aplicación de plaguicidas (Yardim & Edwards, 1998). A pesar de ello, los resultados muestran que hay una diferencia en las especies dominantes entre los años estudiados, siendo especies licósidas las dominantes en 2002 y especies de Araneidae en 2004, sus comportamientos de búsqueda de alimento también son distintos. Estos resultados coinciden con Bostanian (2001), que encontró que la diversidad de arañas en cultivos de frutilla, cortados al ras en su segundo año, estaba representada por las familias Araneidae y Theridiidae. En cambio, en las plantas que no habían sido cortadas, la familia Lycosidae fue la mejor representada.
Si bien en los sistemas agrícolas a menudo es muy difícil determinar qué factores son los que influencian la abundancia y la diversidad de los artrópodos, es probable que una combinación de diferentes prácticas agrícolas determine los ensamblajes y la disponibilidad de los artrópodos en el cultivo (Holland & Luff, 2000). Se puede concluir que la comunidad de artrópodos en un cultivo de frutilla en un mismo predio, varía con los años y con la fenología del cultivo y entre parcelas con plantas con edades diferentes de implantación, aun muy cercanos entre sí. El incremento de la heterogeneidad interna del cultivo, producto de colocar parcelas con plantas de diferentes edades en el mismo predio, puede incrementar la diversidad de la artropodofauna, y se encuentran diferencias en las comunidades en los distintos grupos funcionales considerados. Por ello, y siguiendo lo expresado por Haddad et al. (2001), posiblemente se debería considerar como estrategia para incrementar el impacto de enemigos naturales y su diversidad, tener en cuenta las características estructurales y fenológicas de los cultivos, más que el incrementar el número de especies de plantas cultivables en el predio.
BIBLIOGRAFÍA
1. Altieri, M.A., 1994. Biodiversity and pest management in agroecosystems. Haworth Press, New York, 185 pp. [ Links ]
2. Altieri, M.A., 1999. Agroecología: Bases Científicas para una Agricultura Sustentable. Ed. Nordan-Comunidad. Montevideo-Uruguay, 325 pp. [ Links ]
3. Altieri, M.A. and C. Nicholls, 2004. Biodiversity and pest management in agroecosystems. New York: Haworth Press, 236 pp. [ Links ]
4. Autio, W.; S. Shloeman, J. Clements and W. Coli, 2004. Fruit Program. <http://www.umass.edu/agland/teams/fruit.html. University of Massachusetts Amherst> Consultada el 06/07/2013. [ Links ]
5. Bostanian, N.J., 2001. Spider composition in a renovated and non-renovated strawberry field. Journal of Agricultural and Urban Entomology 18: 201-207. [ Links ]
6. Coll, M.; S. Shakya, I. Shouster and Y. Nenner, 2006. Decision-making tools for Frankliniella occidentalis management in strawberry: consideration of target markets. Entomologia Experamentalis et Applicata 121: 1-9. [ Links ]
7. Colwell, R., 2004. Estimate S. Statistical estimation of species richness and shared species from samples. Version 7.0. <http://viceroy.eeb.uconn.edu/estimates> Consultada el 05/06/2013. [ Links ]
8. Cross, J.; M.A. Easterbrook, A.M. Crook, D. Crook, J. D. Fitzgerald, P.J. Innocenzi, C.N. Jay, and M.G. Solomon, 2001. Review: Natural Enemies and Biocontrol of Pests of Strawberry in Northern and Central Europe. Biocontrol Science and Technology 11(2): 165-216. [ Links ]
9. Fry, G., 1995. Landscape ecology of insect movement in arable ecosystems. In Glen D.M., M.P. Greaves and H. M. Anderson (Eds.): Ecology and integrated farming systems. John Wiley and Sons, Bristol, UK, pp. 177-202. [ Links ]
10. Gomez-Moya, C.A. y F. Ferragut, 2009. Distribución en la planta y eficacia de Noesilius califormicus (Acari: Phytoseiidae) en el control de arañas rojas de cultivos en condiciones de semicampo. Boletín de Sanidad Vegetal Plagas 35: 377-390. [ Links ]
11. Greco, N.M.; G.T. Tetzlaff and G.G. Liljesthrom, 2004. Presence-absence sampling for Tetranychus urticae and its predator Neoseiulus californicus (Acari: Tetranychidae, Phytoseiidae) on strawberry. International Journal of Pest Managment 50 (1): 23-27. [ Links ]
12. Greco, N.M.; N.E. Sánchez and G.G. Liljesthrom, 2005. Neoseiulus californicus (Acari: Phytoseiidae) as a potential control agent of Tetranychus urticae (Acari:Tetranychidae): efecto of pest/predator ratio on pest abundance in strawberry. Experimental and Applied Acarology 37: 57-66. [ Links ]
13. Greco, N.M.; N.E. Sánchez, G.G. Liljesthrom, E. Amnrosio, M.A. Basiglio Cordal, M.F. Cingolani, N. Cluigt, M.F. Gugole y M. Roggiero, 2007. Transferencia de una estrategia de manejo de la "Arañuela roja" a productores de frutilla del cinturón hortícola del Gran La Plata. Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET); Facultad de Ciencias Naturales y Museo (UNLP); Centro de Estudios Parasitológicos y de Vectores (CEPAVE). Página/s: 10. ISSN 1851-877X. <http://www.fcnym.unlp.edu.ar/dependencias/secextension/Experiencia4-2007.pdf> Consultada el 05/04/2013. [ Links ]
14. Haddad, N.M.; D. Tilman, J. Harstad, M. Ritchie and J.M.H Knops, 2001. Contrasting effects of plant richness and composition on insects communities: a field experiment. American Naturalist 158:17-35. [ Links ]
15. Hammer, O.; D.A.T. Harper and P.D. Ryan, 2003. PAST (Paleontological Statistics) version. 1.18. <http://folk.uio.no/ohammer/past> Consultada el 04/05/2013. [ Links ]
16. Holland, J. M. and M. L. Luff, 2000. The effects of agricultural practices on Carabidae in temperate agroecosystems. Integrated Pest Management Reviews 5:109-129. [ Links ]
17. Holling, C. S., 1973. Resilience and Stability of Ecological Systems Annual Review of Ecology and Systematics. Vol. 4: 1-23 (Volume publication date November 1973) DOI: 10.1146/annurev.es.04.110173.000245 [ Links ]
18. Huffaker, C. B. and P.S. Messenger, 1976. Theory and practice of biological control. Academic Press, New York, 788 pp. [ Links ]
19. Krebs, C.J., 1999. Ecological methodology. Second edition. Addison Wesley, Longman, Inc., Menlo Park, California, USA, 620 pp. [ Links ]
20. Laing, J. E., 1969. Life history and life table of Tetranychus urticae Koch. Acarologia 11: 32-42. [ Links ]
21. Lefebvre, M.G.; C. Reguilón, y D.S. Kirschbaum, 2013. Evaluación del efecto de la liberación de Orius insidiosus (Hemiptera: Anthocoridae), como agente de control biológico de trips en el cultivo de frutilla. Revista de Investigaciones Agropecuarias 39 (3): 273-280. [ Links ]
22. Lemme, M.C.; A.P. Jaime de Herrero, D.S. Kirschbaum y A.J. Nasca, 1996. Artrópodos asociados al cultivo de la frutilla, Fragaria x ananassa, en Tucumán, Argentina. Nota de investigación. Vedalia 3: 51-52. [ Links ]
23. Magurran, A., 2004. Measuring Biological Diversity. Blackwell Science Ltd., USA, 256pp [ Links ]
24. McCune, B. and J.B. Grace, 2002. Analysis of Ecological Communities. M & M Software Design, Gleneden Beach, Oregon, USA, 300 pp. [ Links ]
25. Meyke, E., 1999-2004. TAXIS 3.5-Taxonomical Information System. <http://www.bio-tools-net> Consultada el 05/04/2008. [ Links ]
26. Perner, J.; C. Wytrykush, A. Kahmen, N. Buchmann, I. Egerer, S. Creutzburg, N. Odat, V. Audorff and W. Weisser, 2005. Effects of plant diversity, plant productivity and habitat parameters on arthropods abundance in montane European grassland. Ecography 28: 429-442. [ Links ]
27. Pinent, S.M.J.; A. Nondillo, M. Botton, L.R. Redaelli and C.E.C. Pinent, 2011. Species of thrips (Insecta, Thysanoptera) in two strawberry production systems in Rio Grande do Sul State, Brazil. Revista Brasileira de Entomologia 55: 3. [ Links ]
28. Proyecto INTA cartera 2009-2012. Desarrollo de un sistema de producción integrada de frutilla adaptable a las principales cuencas frutilleras argentinas. Coordinador Kirschbaum D. S. <http://inta.gob.ar/proyectos/pnhfa-061281> Consultada el 06/07/2013. [ Links ]
29. Richards, P.C. and J. Schmidt, 1996. The effect of selected dietary supplements on survival and reproduction of Orius insidiosus (Say) (Hemiptera: Anthocoridae). Canadian Entomology 128: 171-176. [ Links ]
30. Ripa, R.; J. Funderbunk, F. Rodiguez, F. Espinosza and L. Mound, 2009. Population Abundance of Frankliniella occidentalis (Thysanoptera: Thripidae) and Natural Enemies on Plant Hosts in Central Chile. Environmental Entomology 38 (2): 333-344. [ Links ]
31. Rondon, S.I.; D.J. Cantliffe and J.F. Price, 2005. The Feeding Behavior of the Bigeyed Bug, Minute Pirate Bug, and Pink Spotted Lady Beetle Relative to Main Strawberry Pests. Environmental Entomology 33(4): 1014-1019. [ Links ]
32. Rondon, S.I.; J.F. Price and D.F. Cantliffe, 2004. Strawberries: Main Pests and Beneficials in Florida. Horticultural Sciences Department, Florida Cooperative Extension Service, Institute of Food and Agricultural Sciences. University of Florida. [ Links ]
33. Saini, E.D.; V. Cervantes y L. Alvarado, 2003. Efecto de la dieta, temperatura y hacinamiento, sobre la fecundidad, fertilidad y longevidad de Orius insidiosus (Say) (Heteroptera: Anthocoridae). Revista de Investigaciones Agropecuarias 32(2): 21-32. [ Links ]
34. Senasa, 2009. <http://www.sinavimo.gov.ar/cultivo/frutilla> Consultada el 06/05/2013. [ Links ]
35. Sorensen, K.A.; Gluber, W.D., Welch, N.C. and Osteen, C., 1997. The importance of pesticides and other pest management practices in U.S. strawberry production. North Carolina Cooperative Extension Service. NAPIAP 1-CA-97. [ Links ]
36. Stumpf, N. and R. Nauen, 2002. Biochemical markers linked to abamectin resistance in Tetranychus urticae (Acari: Tetranychidae). Pesticide Biochemistry and Physiology 72: 111-121. [ Links ]
37. Stumpf, N.; P.W. Claus, W. Kraus, G.D. Moores and R. Nauen, 2001. Resistance to organophosphates and biochemical genotyping of acetylcholinesterases in Tetranychus urticae (Acari: Tetranychidae). Pesticide Biochemistry and Physiology, 69, 131-142. [ Links ]
38. Van Emden, H. F., 1965. The role of uncultivated land in the biology of crop pests and beneficial insects. Scientific Horticulture 17:121-126. [ Links ]
39. Wright, B., 2001. Know Your Friends: Minute Pirate Bugs. <http://www.entomology.wisc.edu/mbcn/kyf101.html> Consultada el 04/03/2013. [ Links ]
40. Yardim, E.N. and C.A. Edwards, 1998. The influence of chemical management of pests, diseases and weeds on pest and predatory arthropods associated with tomatoes. Agriculture, Ecosystems and Environment 70: 31-48. [ Links ]
41. Zalom, F.G.; P.A. Phillips, N.C. Toscano and M. Bolda, 2005. UC IPM Pest Management Guidelines: Strawberry. UC ANR Publication 3468. Statewide IPM Program, Agriculture and Natural Resources, University of California, 76 pp. [ Links ]

