











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
Los ecosistemas acuáticos están formados por una compleja red de interacciones entre factores bióticos y abióticos que acentúa en comunidades de peces Neotropicales debido a su alta diversidad (Oliveira-Fogaça et al. 2003). En este sentido, el estudio de la dieta de peces Neotropicales es un aspecto primordial para entender el funcionamiento de los ecosistemas en los que habitan, así como la función que cumplen en los mismos desde las bases de la malla trófica (Gerking 1994). En esta dinámica trófica, los peces desempeñan un papel crucial al intercambiar materia y energía, lo que sustenta un nicho ecológico en el ecosistema (García-Cardona, Molina y Duque 2021). Este nicho refleja las relaciones existentes entre los ejemplares que componen el sistema (Sánchez-Hernández et al. 2019). Por lo tanto, es crucial comprender la ecología trófica, ya que proporciona información relevante sobre la dinámica y la estructura de la comunidad (Cogua et al. 2013), aportando información para mitigar y contrarrestar los impactos potenciales a los que una comunidad podría estar expuesta (García-Cardona, Molina y Duque 2021).
En estudios de ecología trófica uno de sus objetivos principales es determinar el ítem de mayor importancia alimentaria, lo que provee información fundamental para la descripción de la dieta y hábitos alimenticios de una especie (Hyslop 1980). Esto puede interpretarse tanto en términos de ocurrencia, volumen, peso y/o valor energético y cada uno de estos componentes puede ser analizado en función de la pregunta ecológica que se pretende responder. En la mayoría de los casos, la ocurrencia y cantidad del ítem (dada generalmente por volumen o por peso), se utilizan para resumir su importancia en un contexto ecológico (Bennemann et al. 2006).
El análisis de hábitos tróficos, a través de la evaluación del contenido estomacal, ha sido una práctica frecuente en estudios de tramas tróficas (Amundsen et al.1996). Algunos investigadores consideran que este tipo de estudios permite evaluar aspectos de la ecología alimentaria de las especies en condiciones naturales, lo cual es esencial para el desarrollo de estrategias de manejo de ecosistemas acuáticos (Segatti y Luciana 2003).
El tipo de ambiente y su disponibilidad de alimento condicionan a los peces, ya que pueden tener que elegir entre un hábitat que proporciona una presa más abundante pero más difícil de capturar, y otro con menos presas, pero con menor costo energético final al ser más fáciles de capturar (Crowder y Cooper 1982, Pyke 1984).
Mirande y Koerber (2020), consideran una lista de 570 especies de peces presentes en aguas dulces de Argentina. En el valle aluvial del río Paraná, zona de influencia de la represa de Yacyretá (Argentina), se identificaron 173 especies de peces distribuidas en 11 órdenes y 32 familias (Scipioni et al. 2016), mientras que en el Parque Nacional Pre-Delta, Almirón et al 2008 mencionan 185 especies.
La familia Acestrorhynchidae contiene sólo el género Acestrorhynchus, el cual está representado por 15 especies presentes en una gran variedad de hábitats, todas con preferencias de ambientes lénticos (Saccol-Pereira et al. 2006). Tres especies de Acestrorhynchus (A. bristkii, A. lacustris y A. pantaneiro), son parte de la dieta de otros peces con valor pesquero (Hahn et al. 2000). Al mismo tiempo, estas especies son importantes en la cadena alimentaria debido a su naturaleza piscívora, y podrían desempeñar función crucial en el control de la estructura de la comunidad acuática (Bennemann y Shibatta 2002, Cantanhêde et al. 2008, Rocha 2009, Krinski 2010).
Asimismo, según Krinski (2010), A. pantaneiro es una de las especies dentro del género más reconocida por alimentarse principalmente de peces y en menor cuantía de crustáceos. Sin embargo, se desconocen las especies de peces que constituyen sus presas en estos ambientes, como también, si presenta alguna preferencia. Por tal motivo, el objetivo del presente estudio fue ampliar el conocimiento disponible de A. pantaneiro como especie piscívora con el fin de determinar si tiene preferencia por alguna especie de pez en ambientes del valle aluvial del río Paraná. Para ello, se propuso determinar las especies de peces que componen los distintos ambientes del valle aluvial del río Paraná y comparar la abundancia relativa con la preferencia de especies consumidas.
Materiales y métodos
Selección de sitios de muestreo y medición de variables ambientales. El estudio se llevó a cabo en un tramo de 210 km del río Paraná aguas abajo de la represa de Yacyretá, entre Ituzaingó (ITU) y Puerto González (PGO). Se determinaron cuatro zonas asociadas al canal: 1- ITU, 2- Itá Ibaté (ITA), 3- Puerto Abrá (ABR) y 4- PGO (Figura 1). En cada una se seleccionaron tres sitios representativos del valle aluvial, con conexión permanente con el cauce principal que fluctúa respondiendo a los distintos niveles hidrológicos (Figura 2; Tabla 1).

Figura 1 Mapa de las zonas muestreadas. 1- Ituzaingó, 2- Itá Ibaté, 3- Puerto Abrá y 4- Puerto González.

Figura 2 Fotos representativas de los sitios en cada zona muestreada. A- Ituzaingó (Laguna punta Isla San Martin), B- Itá Ibaté (Laguna Melilla), C-Puerto González (Laguna uno aguas abajo), D- Puerto Abra (Laguna frente al campamento).
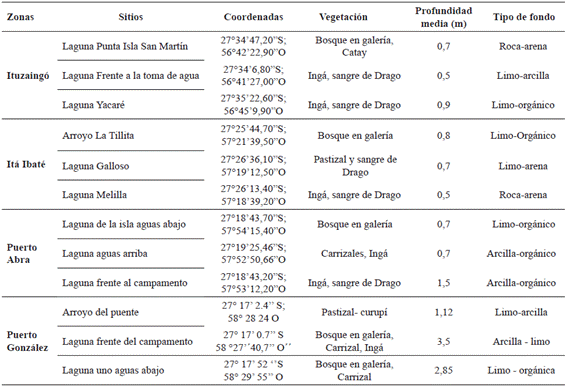
Tabla 1 Características de las zonas, sitios estudiados y de los ambientes del valle aluvial del río Paraná.
Periodicidad del muestreo. El intervalo de tiempo entre cada muestreo fue de 3 a 4 meses, desde mayo de 2016 a febrero de 2018.
Datos ambientales físicos y químicos. En cada fecha de muestreo, se registraron los siguientes parámetros físico-químicos del agua, tales como: temperatura y oxígeno disuelto (YSI®), pH (waterproof), conductividad eléctrica (HANNA®) y transparencia con disco de Secchi. Adicionalmente se estimó la turbidez mediante el uso de un espectrofotómetro portátil (HANNA). Los sitios de muestreo fueron geo-referenciados con un GPS Garmin®.
Obtención de las muestras de peces. Se utilizó un equipo de pesca eléctrica marca Smith Root modelo IV A, montado en una embarcación de fibra de vidrio, a fines de obtener capturas de ejemplares en las áreas de estudio, alimentado con un grupo electrógeno Marca Honda Modelo EP2500CX. El transformador se configuró para rectificar la corriente a corriente continua pulsátil (CCP) entre 60-120 pps a 1000 voltios. El esfuerzo de muestreo fue de 20 minutos resultando en una distancia recorrida, de entre 100 y 200 metros, según la complejidad del ambiente muestreado. Una vez capturados, la eutanasia de los ejemplares fue realizada con una sobredosis de benzocaína (300 mg L-1) (Schroeder et al. 2021). Posteriormente, se fijaron en formol al 10% y luego conservados en alcohol al 70%.
Trabajo en laboratorio. Para cada ejemplar se registró el peso (g), la longitud estándar (mm) y apertura bucal (mm). Se extrajo el estómago por medio de una incisión abdominal y luego se lo pesó y registró en una planilla de cálculos en gramos. Los ítems alimenticios encontrados en los estómagos fueron identificados hasta la mayor determinación taxonómica posible, siguiendo la clasificación de bibliografía disponible (Almirón et al. 2015). Se cuantificó el contenido mediante el recuento de cada ítem, estimando además el volumen de cada uno de ellos utilizando la fórmula de una esfera elipsoide: 4/3πabc; donde a = largo del ítem alimenticio; b = alto y c= porcentaje del ancho (70%). El análisis de los ítems alimenticios fue realizado utilizando una lupa estereoscópica modelo Olympus SZ40.
Análisis de datos. Para el análisis del contenido estomacal se utilizaron los siguientes índices:
Índice de Frecuencia de Ocurrencia (Hyslop 1980): FO: número de estómagos con el ítem A/ número total de estómagos llenos.
Método Volumétrico (Marrero 1994): V: volumen ítem A/volumen de todos los ítems estudiados x 100.
Índice Alimentario (Lauzanne 1975): IA = %V x %FO/100; donde %V es el porcentaje de volumen (V) de cada ítem de la dieta y %FO es el porcentaje de la frecuencia de ocurrencia de un determinado ítem alimenticio.
Para detectar si existen diferencias entre los ítems ingeridos y los disponibles en un gradiente longitudinal del río, se realizó un test estadístico Chi X2 con la hipótesis de que A. panteneiro se alimenta de las especies más abundantes en los ambientes estudiados. Para eso se dividió en dos grandes áreas: un área cercana a la represa (ITU- ITA) y un área más lejana (ABRA-PGO). Cabe destacar que, para esta estadística se tomaron en cuenta solo las especies identificadas, dejando de lado el gran porcentaje de ítems digeridos que no se pudo identificar. Asimismo, para buscar una correlación entre el tamaño de la boca y del ítem alimenticio, se realizó una regresión lineal mediante el uso de planillas de cálculos Excel, entre los valores de largo y ancho de la boca con el volumen de los ítems consumidos por A. pantaneiro.
Resultados
Al final del período de estudio se capturaron 140 ejemplares de A. pantaneiro, los que presentaron una longitud estándar entre 27 y 240 mm (media = 98,05 ± 55,28 mm). De estos, 59 ejemplares presentaron estómagos vacíos (42%), mientras que 81 ejemplares presentaron contenidos estomacales (58%). En estos últimos, en 69 ejemplares se observó un solo tipo de ítem alimentario y el resto más de un tipo. Los ítems registrados fueron clasificados en: Cichliformes (varias especies A, B), Characiformes (varias especies C, D, E, F, G, H, I, J, K), Siluriformes (1 especie, L), peces digeridos (M) y vegetales superiores (N). Debido al proceso de digestión, algunos ítems no pudieron ser identificados hasta el nivel taxonómico de especie o familia.
El análisis de los contenidos estomacales mostró que los principales ítems que componen la dieta de A. pantaneiro fueron peces. La mayor parte de estas especies no han podido ser determinadas (agrupadas como peces digeridos), en segundo orden se situaron especies del género Crenicichla (Cichliformes) y en el tercer orden se ubicaron las especies identificadas dentro del orden Characiformes (Tabla 2). En la Figura 3 se observa el porcentaje del Índice Alimentario calculado.

En las cuatro zonas, A. pantaneiro fue capturado junto a otras especies de peces, siendo las más abundantes Moenkhausia bonita, Poptella paraguayensis, Aphyocharax dentatus, Otocinclus vestitus, entre otras. El análisis de la prueba de Chi cuadrado reveló que existen diferencias significativas (p<0,05) entre las proporciones estimadas de los ítems ingeridos y los disponibles, como se muestra en la Figura 4. Por otro lado, los valores de R2 para la correlación entre el tamaño de la presa y el largo (R2: 0,1459) y ancho (R2: 0, 2161) de la boca de la especie en estudio no demostraron una relación significativa.

En la Figura 5 se observan las abundancias relativas de algunas especies disponibles en el valle aluvial durante el periodo de muestreo en cada uno de los sitios. A la derecha del gráfico puede observar que el género Crenicichla no es el más abundante.
Temporalmente no se observaron diferencias, por lo cual no se hace mención en este estudio.

Discusión
Los resultados del presente estudio muestran que A. pantaneiro es piscívoro coincidiendo con otros estudios, tanto en juveniles como adultos (Catella y Petrere-Junior 1996, Resende et al. 1996, Lowe-Mcconnell 1999, Machado 2003, Britski et al. 2007). Krinski (2010), menciona que en el Pantanal de Mato Grosso (Brasil), esta especie, además de ser piscívora, se alimenta de camarones (Crustacea) indicando plasticidad trófica; como así también mostró un comportamiento de canibalismo, características que no se presentaron en este estudio. También, cabe mencionar la identificación de material vegetal en la dieta de algunos ejemplares observado en este estudio, que no fue mencionado por otros autores para A. pantaneiro, pero si para A. lacustris en el río Tibagi (Bennemann 1996). Esta ingesta podría deberse a una ingestión accidental, relacionada a captura de sus presas mediante persecución tal como fue mencionado por Krinski (2010) en Brasil.
Un porcentaje considerable de estómagos de A. pantaneiro colectados se encontraron vacíos, tal como fue reportado por Krinski (2010), para A. pantaneiro, A. lacustris y A. britskii. Según Gerking (1994), esta incidencia de estómagos vacíos es una característica de especies carnívoras; puesto que la saciedad en estos peces ocurre en un periodo más largo, lo que determina esa alta frecuencia de estómagos vacíos (Zavala- Camin 1996).
Según Gerking (1994), en general, los peces tienen la habilidad de adecuarse a una gran variedad de fuentes de alimento. Esta habilidad de sacar provecho a cada fuente de alimento, en un determinado período, es denominada plasticidad trófica, partiendo del principio de que los peces son lo suficientemente flexibles para cambiar de un recurso alimentario a otro en situaciones de escasez. Los resultados de este trabajo sugieren que esta especie es considerada generalista por la gran variedad de tipos de presas consumidas. Sin embargo, en estos ambientes, muestra cierta preferencia por el género Crenicichla con altos valores en algunos índices en cuanto a las especies identificadas, resaltando que este género no se presenta como una de las especies más abundantes en el valle aluvial, tal como fue mencionado por Scipioni et al. (2016).
Dill (1983), comenta que la flexibilidad alimentaria es una característica adaptativa importante del comportamiento alimentario, una vez que los ambientes naturales varían espacial o temporalmente, los peces responden a la baja disponibilidad alimentaria alterando su comportamiento. En relación con el gradiente ambiental estudiado, A. pantaneiro no presentó una variación como expresan Díaz-Pardo y Trujillo (1995), opinando que los diferentes organismos, que sirven de alimento para los peces, sufren fluctuaciones determinadas por la dinámica ambiental.
Según Mittelbach y Persson (1998), el tamaño de la presa consumida varía en función del tamaño del depredador; depredadores más pequeños tienden a consumir presas más pequeñas, mientras que los depredadores más grandes pueden consumir presas de diferentes tamaños. Krinski (2010) observó, que los especímenes más grandes de Acestrorhynchus solo consumieron presas más grandes. En nuestro estudio, no encontramos una diferencia significativa en el tamaño de la presa con relación al tamaño del depredador; el dientudo consumió presas de diferentes tamaños, lo que sugiere que no hay una correlación clara. Esto refuerza la idea de que A. pantaneiro, entre las opciones disponibles, no busca específicamente presas más grandes para consumir. Tampoco muestra preferencia por las más abundantes, capturando de manera diferencial ejemplares de Crenicichla.
La información obtenida en este estudio confirma que A. pantaneiro se comporta, en ambientes asociados del valle aluvial del río Paraná, como una especie piscívora y selectiva a la hora de capturar sus presas, enriqueciendo el conocimiento sobre la ecología trófica de esta especie en dicho sitio. Además, se observa una tendencia hacia el consumo de especies del género Crenicichla en la dieta de A. pantaneiro en el valle aluvial del río Paraná, específicamente en la zona comprendida entre la represa de Yacyretá y la confluencia con el río Paraguay.

