










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMultequina
versión On-line ISSN 1852-7329
Multequina vol.9 no.1 Mendoza 2000
ARTICULO ORIGINAL
Torneutes pallidipennis reich (Coleoptera: Cerambycidae) en algarrobos negros del Chaco Arido de Córdoba. Efecto de la condición de sitio en el grado de infestación
Torneutes pallidipennis reich (Coleoptera: Cerambycidae) in the region of the Arid chaco of Córdoba. Effect of the site condition in the infestation on black algarrobo trees
Ana A. Córdoba1, V. M. Azzuferi1, R.O. Coirini1, U. Karlin1 y F. Casanoves1
1 Facultad de Ciencias Agropecuarias, Universidad Nacional de Córdoba, Argentina. C.C. 509, 5000 Córdoba - Argentina (E-mail: mam@agro.uncor.edu)
RESUMEN
En la región del Chaco Árido de Córdoba, Argentina, el recurso forestal se encuentra degradado debido al manejo poco racional y a la acción de insectos xilófagos. Torneutes pallidipennis Reich, ha sido señalado como una de las principales especies xilófagas de la zona. Cuatro sitios, con dos repeticiones cada uno, fueron utilizados para determinar el grado de influencia de la condición de sitio en la infestación de T. Pallidipennis sobre Prosopis aff. flexuosa. Los sitios seleccionados surgen de un arreglo factorial de la biomasa herbácea y densidad arbórea, con dos niveles cada uno. Además se relacionó el grado de infestación con el diámetro de los árboles. No se detectó interacción entre los factores biomasa herbácea y densidad arbórea, tampoco diferencias significativas entre la variable daño y densidad arbórea, mientras que el ANAVA arrojó diferencias significativas entre la variable daño y el factor biomasa herbácea, existiendo una relación inversa. Además se detectó una fuerte dependencia directa entre el diámetro de los árboles y el daño, con una marcada diferencia en la infestación entre árboles menores y mayores a 15 cm de DAP.
Palabras clave: Coleoptera; Xilófago; Sitio; Infestación
SUMMARY
In the region of the Arid Chaco of Córdoba, Argentina, the forest resource is degraded due to low rational management and to the action of xylophagous insects. Torneutes pallidipennis Reich, has been indicated as one of the main xylophagous species of the zone. Four sites, with two repetitions each one, were used to determine the effect of the condition of site in the infestation of T. pallidipennis on Prosopis aff. flexuosa. The selected sites arise from a factorial adjustment of the factors grass biomass and tree density, with two levels each one. In addition, the infestation degree was related to the diameter of the tree. Interaction between the grass biomass and arboreal density was not detected, either significant differences between the variable damage and arboreal density, whereas the ANAVA threw significant differences between the variable damage and the factor grass biomass, existing an inverse relation. In addition a strong direct dependency was detected between the diameter of the tree and the variable damage, with an important difference in the infestation between trees lower and higher than 15 cm in breast diameter.
Key words: Coleoptera; Xylofagous; Site; Infestation.
INTRODUCCIÓN
En la región fitogeográfica del Chaco Árido de Córdoba los «algarrobos negros», Prosopis aff. flexuosa (Palacios y Bravo, 1981) , aportan el 70% del recurso forestal (Karlin et al., 1992). A pesar de su importancia, el manejo poco racional y la acción de insectos xilófagos han llevado a la degradación a los bosques nativos. En elaño 1988 Mazzuferi y Coirini identificaron a Torneutes pallidipennis Reich (Coleoptera: Cerambycidae) como el principal agente causal de los daños producidos en «algarrobos negros» en dicha región.
Las larvas de T. pallidipennis realizan galerías longitudinales descendentes ubicadas en el duramen que llegan a las ramas de segundo y tercer orden (Di Iorio, 1996). Estas, al igual que todos los Torneutini, han adoptado una estrategia de vida especial por estar adaptadas a vivir en tejidos muertos, pero en plantas vivas con un duramen diferenciado (Di Iorio, 1996). Este comportamiento los diferencia de los otros Cerambicidos que infestan solo árboles muertos, secos o debilitados (Nielsen, 1981). Sin embargo la infestación de T. pallidipennis parece ser más severa en sitios disturbados. Una de las formas de determinar el estado de degradación de un sitio es a través de la presencia de suelo descubierto, porcentaje de gramíneas anuales (Karlin et al., 1994) y la biomasa herbácea (Karlin et al., 1992; Echaves et al., 1998). Estos, junto con la densidad arbórea, es lo que a continuación se denomina «condición de sitio». Echaves et al. (1998) determinaron que la infestación deMegacyllene robiniae Forster (Coleoptera: Cerambycidae), en árboles vivos de Robinia pseudoacacia (Hopkins, 1907; Hall, 1942; Berry, 1945; Harman et al., 1985 y Echaves et al., 1998) es mayor en los sitios más pobres, donde la biomasa herbácea es menor.
El número de árboles por superficie también puede predisponer a un árbol a ser infestado. Hilje et al. (1991), Arguedas-Gamboa y Quiróz-Rodriguez (1997) sostienen que cuando los árboles se encuentran en alta densidad se produce mayor competencia, lo que los hace más propensos al ataque de plagas. Por el contrario, Craighead (1950), Metcalf and Flint (1965) y Brugnoni (1980) sostienen que los árboles en plantaciones abiertas están más sujetos al ataque de Cerambícidos.
Otro factor relacionado con la infestación de este insecto es el diámetro del árbol. Mazzuferi y Coirini (1988) junto con Fiorentino y Diodato de Medina (1988, 1995) coinciden en señalar que la frecuencia de árboles infestados por T. pallidipennis se manifiesta en forma creciente según aumenta el DAP (diámetro a la altura del pecho) habiendo un importante incremento de árboles infestados a partir de los 12,5-15 cm de DAP según Fiorentino y Diodato de Medina (1988, 1995). En ambos estudios resultaron más frecuentes los árboles con el 100% de las ramas infestadas, seguidos de aquellos sin infestación.
El objetivo de este estudio fue determinar el grado de influencia de la condición de sitio en la infestación de T. pallidipennis sobre «algarrobos negros» en el Chaco Árido de Córdoba y relacionar el grado de infestación de dicho insecto con el diámetro de los árboles.
MATERIAL Y MÉTODO
El estudio se llevó a cabo en la localidad de Chancaní ubicada a 31º 25' latitud sur y 65º 26' latitud oeste, Departamento Pocho, Provincia de Córdoba, Argentina, respondiendo a la clasificación fitogeográfica de Chaco Árido según Morello et al. (1985).
Se definieron cuatro condiciones de sitio (C1, C2, C3 y C4) que surgen de un arreglo factorial de la biomasa herbácea (BH) y densidad arbórea (DEN) con dos niveles cada uno (Tabla 1).
Tabla 1. Valores de densidad arbórea y biomasa herbácea promedio de cada condición de sitio y su respectivo valor medio de infestación
Table 1. Values of arboreal density and herbaceous biomass average of each condition of site and its respective average value of infestación
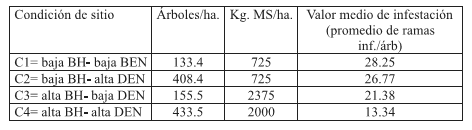
En cada uno de los sitios, la densidad arbórea se determinó realizando 2 transectas en fajas al azar de 50m x 6m. La presencia de suelo descubierto y gramíneas anuales se determinó recorriendo los sitios, mientras que la estimación de biomasa herbácea se llevó a cabo mediante el método de doble muestreo (Díaz, 1992), realizando 2 transectas por sitio con 50 cuadrados de estimación visual cada una.
Se seleccionaron, de forma independiente, dos sitios de cada una de las categorías a modo de repetición. En cada uno, la toma de datos se realizó sobre 50 árboles (Prosopis flexuosa) situados en un itinerario concéntrico, que determinó el tamaño de las parcelas de estudio. Se consideró como «árbol» a aquellos individuos con diámetro de tronco >5 cm de DAP (Naveiro et al., 1999). Los muestreos se efectuaron entre diciembre de 1999 y febrero de 2000, debido a que los adultos de T. pallidipennis emergen durante el período estival (Di Iorio, com. pers.).
Con el fin de evaluar el efecto de la condición de sitio sobre el grado de infestación, se determinó el grado de infestación (porcentaje de ramas infestadas) de los árboles en base a la sintomatología que presentaban (Arguedas-Gamboa y Quiróz-Rodriguez, 1997); para ello se calculó el número de ramas infestadas (de segundo y tercer orden) respecto al total de ramas del árbol. Para determinar si una rama estaba infestada, se tuvo en cuenta la presencia-ausencia de orificios de salida de adultos, antiguos o recientes (Naveiro et al., 1999), cuyo reconocimiento se llevó a cabo siguiendo las descripciones de Fiorentino y Diodato de Medina (1988, 1995) y Di Iorio (1996). Con los datos obtenidos se realizó un Análisis de la Varianza para la variable daño con los factores BH, DEN y la interacción BH*DEN, y una distribución de frecuencias del grado de infestación para cada condición de sitio.
Para relacionar el grado de infestación con el diámetro del árbol, se estimó el DAP (a 1,3 m de la base aproximadamente) y se construyeron intervalos de clases a partir de los 5 cm (Fiorentino y Diodato de Medina, 1988, 1995) con una amplitud de 10 cm entre clases diamétricas (DAP<15; DAP15-25; DAP25-35 y DAP>35 cm). En base al grado de infestación de los árboles, se realizó una tipificación por clase de infestación (Fiorentino y Diodato de Medina, 1988, 1995; Mazzuferi y Coirini, 1988): Infestación clase A: 100-75% de las ramas dañadas, clase B: 75-50%, clase C: 50- 25%, clase D: 25% o menos, clase E: sin ramas dañadas. Los datos fueron analizados mediante una tabla de contingencia usando el estadístico Chi-cuadrado. Además, se analizó la distribución de frecuencias del grado de infestación en relación a las clases diamétricas Total (sin distinción de sitio), y para cada condición de sitio.
RESULTADOS
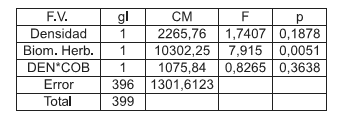
El análisis de la varianza para la variable daño no mostró interacción significativa entre los factores BH*DEN (p=0,3638) como así tampoco diferencias significativas con el factor DEN (p=0.1878); mientras que el ANAVA arrojó diferencias significativas para el factor BH (p=0.0051) (Tabla 2).
Tabla 2. Análisis de la Varianza donde se evalúa el efecto de la interacción de los factores BH*DEN, factor BH y factor DEN sobre la variable daño, con p=0.05
Table 2. Analyses of the Variance where the effect of the interaction of factors BH*DEN, factor BH and factor GIVE on the variable damage is evaluated, with p=0.05.
Como se observa en la Figura 1, en todas las condiciones de sitio existe un gran predominio de árboles sanos con una tendencia generalizada al aumento en la frecuencia en las condiciones de alta BH.
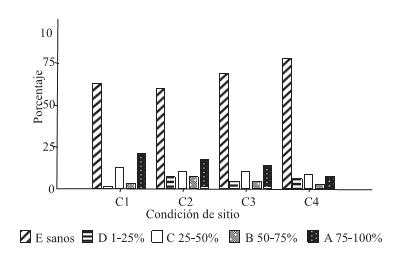
Figura 1. Distribución de frecuencias de las clases de infestación para cada condición de sitio (C1, C2, C3 y C4)
Figure 1. Frequency allocation of the classes of infestación for each condition of site (C1, C2, C3 andC4)
Mientras que las clases de infestación A y C presentan mayor frecuencia en la condiciones de sitio de alta BH. Los resultados obtenidos arrojan una relación inversa entre el factor BH y el valor medio de infestación, es decir que en sitios de baja BH (C1 y C2) la infestación es mayor respecto a aquellos de alta BH (C3 y C4) (Tabla 1).
Referido al grado de infestación de T. pallidipennis en relación a las clases diamétricas, el análisis de distribución de frecuencias de la clase de infestación en relación a las clases diamétricas Total (N=400) y para cada condición de sitio (C1, C2, C3 Y C4), arrojó diferencias significativas entre árboles mayores y menores de 15cm de DAP. Asimismo, el análisis de distribución de frecuencias entre las clases diamétricas de árboles con DAP>15 cm (DAP 15-25, 25-35 y >35cm) no arrojó diferencias significativas para las condiciones C1 y C4 (Tabla 3).
Tabla 3. Frecuencias absolutas y valores de probabilidad de clase de infestación en relación a las clases diamétricas
Table 3. Absolute frequencies and probability values by infestation class in relation to the diametric classes
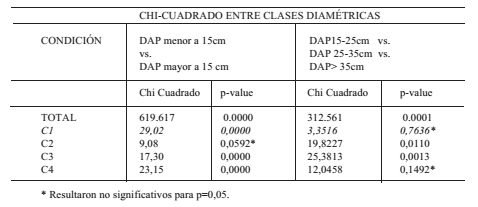
El análisis de la distribución de frecuencia de árboles sanos e infestados por clase diamétrica arrojó como resultado que existe una relación directa entre la infestación y el DAP. De los árboles con DAP>15 cm sólo el 15.27% estaban infestados, mientras que el 100% de los árboles de DAP>35cm presentaban signos de infestación.
De los 400 árboles analizados el 33,5% estaban infestados, de los cuales el 44% presentaba infestación Clase A, el 12% Clase B, el 30,6% clase C y el 13,4% Clase D. En la Figura 2 se puede observar que la frecuencia de infestación Clase A comienza a partir de la primer clase diamétrica, aumenta gradualmente con el incremento del diámetro, y en la última clase diamétrica (DAP>35cm) se incrementa en más del doble constituyendo el 62,5% de los árboles.
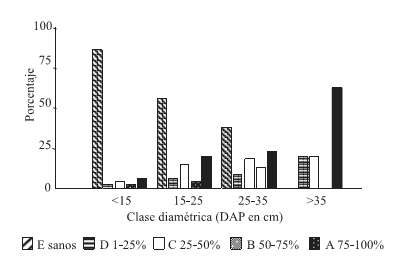
Figura 2. Distribución de frecuencias de las clases de infestación por clases diamétricas, sin distinción de sitios, TOTAL
Figure 2. Frequency allocation of the classes of infestación for diametric classes, without distinction of sites, TOTAL
La frecuencia de infestación Clase A resultó la más alta cualquiera fuese la clase diamétrica.
En las condiciones de sitio C1 y C2 (Figuras 3 y 4 ) se obtuvo una alta frecuencia de árboles infestados en las clases diamétricas menores (DAP<15 y 15- 25 cm).
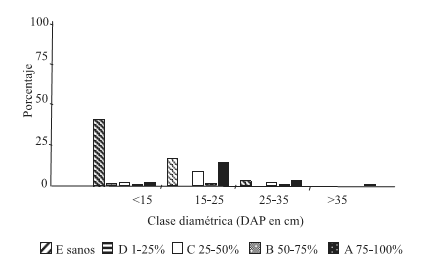
Figura 3. Distribución de frecuencia de las clases de infestación por clases diamétricas, condición de sitio bajaBH-bajaDEN (C1)
Figure 3. Frequency allocation of the classes of infestación for diametric classes, condition of site low HB – lowDEN (C1)
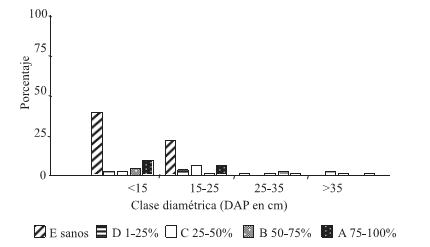
Figura 4. Distribución de frecuencia de las clases de infestación por clases diamétricas, condición de sitio bajaBH-altaDEN (C2)
Figure 4. Frequency allocation of the classes of infestación for diametric classes, condition of site low HB – high DEN (C2)
Para la condición de sitio C1, la mayor frecuencia de árboles infestados se encuentra en la clase diamétrica DAP 15- 25cm, con similar proporción de árboles sanos y con infestación Clase A. Dicha clase de infestación resultó ser predominante, cualquiera fuera la clase diamétrica. En la condición de sitio C2, el mayor porcentaje de árboles infestados corresponde a las clases diamétricas DAP<15 (con un alto porcentaje de infestación clase A) y 15-25cm.
En las condiciones de sitio C3 y C4 (Figuras 5 y 6) la frecuencia de las distin tas clases de infestación resultaron bajas, no superando el 7% en C3 y el 4% en C4.
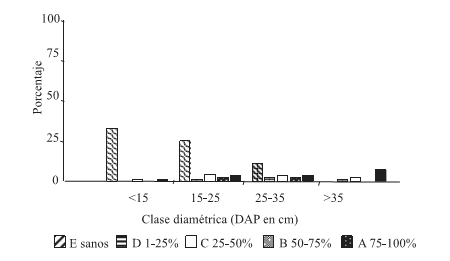
Figura 5. Distribución de frecuencia de las clases de infestación por clases diamétricas, condición de sitio altaBH-baja DEN (C3)
Figure 5. Frequency allocation of the classes of infestación for diametric classes, condition of site high HB –low DEN (C3)
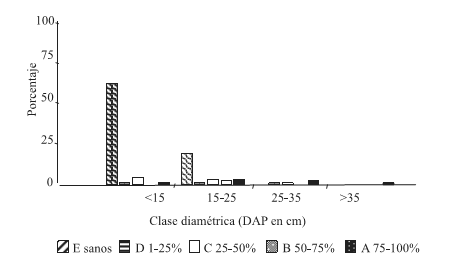
Figura 6. Distribución de frecuencia de las clases de infestación por clases diamétricas, condición de sitio altaBH-alta DEN (C4)
Figure 6. Frequency allocation of the classes of infestación for diametric classes, condition of site high HB – high DEN (C4)
En ambas condiciones, la infestación Clase A resultó con mayor frecuencia en las clases diamétricas mayores (DAP 25-35 y >35cm).
En la condición C3, se observa una tendencia al aumento de la infestación hacia las clases diamétricas mayores. La frecuencia de árboles sanos disminuye paulatinamente con el incremento del diámetro. Mientras que en C4 se obtuvo una disminución de tres veces en la frecuencia de árboles sanos de la clase diamétrica DAP<15 cm a la siguiente clase a partir de la cual la frecuencia se hace cero.
DISCUSIÓN
Existe efecto de la condición de sitio en la infestación de T. pallidipennis, pero esta se debe solamente a la calidad del sitio y no a la densidad, disminuyendo en sitios de mayor calidad, de alta biomasa herbá- cea (alta BH) tanto la frecuencia de árboles infestados como el grado de infestación. Esto concuerda con lo establecido por Hopkins (1907), Hall (1942), Berry (1945), Harman et al. (1985) y Echaves et al. (1998) quienes determinaron que el daño producido por M. robiniae en R. pseudoacacia es mayor en los sitios más pobres y disturbados (con baja BH). A diferencia de lo establecido por Craighead (1950), Metcalf and Flint (1965) y Brugnoni (1980), no se encontró efecto del factor densidad arbórea (entre 133,4 y 433,5 árb/ha) en la infestación de T. pallidipennis. Estos resultados concuerdan con observaciones de Di Iorio, quien sostiene que los árboles pueden estar infestados o no, independientemente de la densidad de los árboles del lugar (Di Iorio com. pers.).
Al igual que Fiorentino y Diodato de Medina (1988, 1995) y Mazzuferi y Coirini (1988), se determinó que existe una relación directa entre la frecuencia de árboles infestados y el DAP, con una marcada diferencia en la infestación de árboles menores y mayores de 15 cm DAP en los que la infestación es baja, con un alto porcentaje de árboles sanos; mientras que a medida que el DAP se incrementa desde los 15cm, la infestación aumenta, llegando a estar infestados el 100% de los árboles mayores a 35 cm. Esto podría estar relacionado con las propiedades de la madera que se encuentran asociadas con el DAP. Martinez et al. (1998), determinaron que a partir de los 15-20 cm de DAP, los «algarrobos negros» presentan igual proporción de albura y duramen, como así también las mismas propiedades fisico-mecánicas, que los árboles de la misma especie y de mayor diámetro. Por ello, la mayor infestación en individuos de diámetros mayores podría deberse a un aumento en el número de episodios de infestación que han sufrido los árboles más viejos.
El porcentaje de árboles infestados resultó ser la mitad de lo establecido por Mazzuferi y Coirini (1988). Esto podría ser explicado por el predominio que presentan en la actualidad «algarrobos negros» con diámetros menor a 15 cm de DAP, entre los cuales predominan los árboles sanos, sumado a la importante disminución debido a la tala que han tenido los árboles de diámetros mayores (Karlin et al., 1992), que presentan mayor infestación.
De acuerdo con Mazzuferi y Coirini (1988) y Fiorentino y Diodato de Medina (1988, 1995), son de mayor frecuencia los árboles con infestación clase A para todas las clases diamétricas, incluso para la clase DAP<15cm. Esto podría deberse a que los árboles infestados son más propensos a ser reinfestados (Di Iorio com. pers.).
CONCLUSIONES
- Existe efecto de la condición de sitio en la infestación de T. pallidipennis, pero ésta se debe solamente a la calidad del sitio y no a la densidad arbórea, disminuyendo en sitios de mayor calidad tanto la frecuencia de árboles infestados como el grado de infestación.
- En sitios de menor calidad, hay una tendencia generalizada hacia un aumento en la frecuencia y grado de infestación de árboles de diámetros menores de 25cm.
- Existe una relación directa entre la frecuencia de árboles infestados y el DAP.
A la luz de estos resultados, y dada la importancia del recurso forestal algarrobo en la zona de estudio, consideramos de importancia realizar estudios futuros sobre la cuantificación de la pérdida económica producida por la infestación de T. pallidipennis, como así también la implementación de manejo fitosanitario adecuado del monte en el corto plazo, tendiente a disminuir los daños producidos por este insecto.
AGRADECIMIENTOS
Los autores agradecen a los productores de Chancaní, N. Ceballos, F. Machuca, T. Sanchez y J. Gonzalez. También a J. M. Díaz, A. Von Müller, E. Biurrun , M. Karlin y M. Guerra, que colaboraron en el trabajo de campo, al Dr. O. Di Iorio y la Biól. L. Pagliari por el aporte de sus comunicaciones personales y críticas al trabajo.
BIBLIOGRAFÍA
1. ARGUEDAS-GAMBOA, M. y L. QUIRÓSRODRIGUEZ, 1997. Experiencias y perspectivas del manejo de plagas forestales en Costa Rica. Manejo Integrado de Plagas (Costa Rica). 45:34-42. [ Links ]
2. BERRY, F. H., 1945. Effect of site and locust borer on plantations of black locust in the Duke Forest. Journal Forestry. 43 (10): 17-54. [ Links ]
3. BRUGNONI, H.C., 1980. Plagas forestales, zoofitófagos que atacan las principales especies forestales naturales y cultivadas de la República Argentina. Ed. Hemisferio Sur S.A. Primera Edición, 107-175. [ Links ]
4. CRAIGHEAD, F. G., 1950. Insects Enemies of Eastern Forests. U.S. Department of Agriculture, Miscellaneous Publication. 657: 2-271. [ Links ]
5. DÍAZ, R. O., 1992. Evaluación de los recursos forrajeros del Chaco Árido,. En U. O Karlin, y O. R Coirini (eds): Sistemas Agroforestales Para Pequeños Productores De Zonas Áridas,. Proyecto GTZ. Desarrollo agroforestal en comunidades rurales del Noroeste Argentino, Facultad de Ciencias Agropecuarias, Universidad Nacional de Córdoba. p.19-23 [ Links ]
6. DI IORIO, O. R., 1996. Torneutini (Coleóptera: Cerambycidae) of Argentina. Part 1. Their relation to the «South American area of Prosopis L.» and to the xerophilous Acacia Miller (Mimosaceae). G. it. Ent. 8: 125-155. [ Links ]
7. ECHAVES, V.D., D.M. HARMAN and A.L. HARMAN, 1998. Site quality in relation to damage by locust borer, Megacyllene Robiniae Forster in Black Locust. Journal of Entomological Science. 33 (1) 106-112. [ Links ]
8. FIORENTINO, D. C. y L. DIODATO DE MEDINA, 1988. Torneutes pallidipennis Reich 1837 (Coleoptera: Cerambycidae) taladro de Prosopis. Actas IV Congreso Forestal Argentino. Santiago del Estero, Argentina. 3: 755-760. [ Links ]
9. FIORENTINO, D. C. y L. DIODATO DE MEDINA, 1995. Torneutes pallidipennis Reich 1837 (Coleoptera: Cerambycidae) taladro de Prosopis. Resúmenes Taller Nacional de Prosopis. Universidad Nacional de Santiago del Estero, p. 755-760. [ Links ]
10. HALL, R. C., 1942. Control of the locust borer. U.S.D.A., 626: 9. [ Links ]
11. HARMAN, D. M., M. A. VAN TYNE and W. A. THOMPSON, 1985. Comparison of locust borer attack on coal stripmined lands and lands not mined. Ann. Entomol. of Soc. Amer. 78: 50-53. [ Links ]
12. HILJE, L., M. VIQUEZ, C. ARAYA y F. SCORZA, 1991. El manejo de enfermedades y las plagas forestales en Costa Rica. Manejo integrado de plagas forestales. 19 : 34-39. [ Links ]
13. HOPKINS, A. D., 1907. The locust borer and methods for its control. U.S.D.A. Bureau of Entomol. Circ, 83: 8. [ Links ]
14. KARLIN,U.O, R.O. COIRINI, L. PIETRARELLI y E. PERPIÑAL, 1992. Caracterización del Chaco Árido y propuesta de recuperación del recurso forestal. En: KARLIN, U. O y R. O. Coirini, 1992: Sistemas agroforestales para pequeños productores de zonas áridas, Proyecto GTZ. Desarrollo agroforestal en comunidades rurales del Noroeste Argentino, p. 7-12 Facultad de Ciencias Agropecuarias, Universidad Nacional de Córdoba, 104 p. [ Links ]
15. KARLIN, U. O., L. A. CATALÁN y R. O. COIRINI, 1994. Cuáles son los «renovables» recursos naturales del Chaco?. En: La naturaleza y el hombre en el Chaco Seco, Proyecto GTZ. Desarrollo agroforestal en comunidades rurales del Noroeste Argentino, Facultad de Ciencias Agropecuarias, Universidad Nacional de Córdoba. 3: 47-75. [ Links ]
16. MARTINEZ R. H., G. MORENO y E. PAN, 1998. Posibilidades de utilización de árboles de algarrobos negros de pequeños diámetros. Actas (CD) del primer congreso latinoamericano del UFRO en Valdivia, Chile. 144 p. [ Links ]
17. MAZZUFERI, V. y R. O. COIRINI, 1989. Análisis del daño producido por taladros en el Chaco Árido de Córdoba. Jornadas técnicas de Uso múltiple del bosque y sistemas agroforestales. El Dorado, Misiones. 15 p. [ Links ]
18. METCALF, C. L. and W. P. FLINT, 1965. Insectos destructivos e insectos útiles, sus costumbres y su control. Compañía Editorial Continental S.A. Calzada de Tlapan Nº 4620. México 22. D.F. 216 p. [ Links ]
19. MORELLO, J., L. PROTOMASTRO, SANCHO LUZ y C. BLANCO, 1985. Estudio macroecológico de los Llanos de La Rioja. Serie del cincuentenario en la Administración de Parques Nacionales. 5: 1-53. [ Links ]
20. NAVEIRO, F., F. N. PULIDO, J. D. DEL POZO, A. MORCUENDE, M. A. GONZÁLEZ y J. MUÑÓZ, 1999. Situación fitosanitaria del arbolado de las dehesas en Extremadura: Influencia de las labores silvícolas. Boletín de Sanidad Vegetal, Plagas. 25: 425-433. [ Links ]
21. NIELSEN, D. G., 1981. Studying biology and control of borer of woody plants. Bull. Entomol. Soc. Amer. 27 (4): 251-257. [ Links ]
Recibido: 07/2000
Aceptado: 11/2000

