











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCIÓN
Las comunidades biológicas difícilmente es capan al accionar de las actividades humanas. Los estudios que consideran al uso de suelo como parte de las fuerzas que moldean y determinan la composición de las comunidades biológicas son cada vez más frecuentes (Karp et al., 2018; Jakovac et al., 2021; Romdhane et al., 2022). Los hongos liquenizados, organismos simbióticos complejos que consisten en asociaciones entre hongos (micobiontes) y algas o cianobacterias (fotobiontes) (Grube & Berg, 2009), forman parte de las comunidades más afectadas por los ambios en el uso del suelo. Existen numerosos estudios que relacionan la composición de comu nidades de hongos liquenizados con diferentes tipos de usos de suelo (Stofer et al., 2006; Pinho et al., 2012; Chuquimarca et al., 2019; Filippini et al., 2020; Rai et al., 2021). Los diferentes tipos de usos del suelo generan distintos efectos sobre las comunidades de hongos liquenizados. Según Zarabska-Bożejewicz & Kujawa (2018) los pai sajes agropecuarios pueden afectar la diversidad funcional de hongos liquenizados. Por otro lado, en áreas de uso agropecuario en la República Argentina, podemos encontrar especies que son severamente afectadas por el uso de agroquími cos, tales como Usnea amblyoclada (Carrera & Carreras, 2011) mientras que otras resultan tole rantes a estas condiciones (Filippini et al., 2020), como tal es el caso de Ramalina celastri (Krog & Swinscow, 1976).
Los hongos liquenizados pueden ser una va liosa herramienta para monitorizar los cambios ambientales en los entornos urbanos (Koch et al., 2019). En este sentido, se han encontrado diferencias sobre los rasgos del fotobionte y la forma de crecimiento del hongo liquenizado en respuesta a la densidad de las urbanizaciones (Koch et al., 2019). En la Argentina existen nu merosos trabajos en los que se han estudiado las comunidades de hongos liquenizados en relación con las urbanizaciones y sus cambios asociados (e.g., González et al., 1998; Carreras et al., 2005; Mateos & Gonzalez 2016; Marié et al., 2016).
El partido de Luján, en la provincia de Buenos Aires, está conformado por pequeñas lo calidades emplazadas en una vasta matriz agro pecuaria. En base a las diferentes intervenciones y acciones en el territorio, podemos encontrar en líneas generales tres tipos de uso del suelo: Agropecuario o Agrícola, Residencial y Urbano (Código de Ordenamiento Urbano, 2019). Esta caracterización ha permitido la exploración de la asociación de los diferentes usos del suelo con la biodiversidad local (Di Ciocco et al., 2014; Gantes et al., 2014; Sarti et al., 2022). Particularmente para dicho partido, existe una única referencia sobre la composición de la comunidad de lique nes locales (Bardelás, 2012) para la cual se regis traron siete especies de líquenes cortícolas. Por lo tanto, hasta el momento, las comunidades de hongos liquenizados no han sido exploradas en profundidad y, particularmente, no existen tra bajos que las relacionen con los diferentes usos del suelo.
En este marco, el objetivo del presente traba jo es identificar las especies de hongos liqueniza dos asociadas a diferente uso del suelo dentro del territorio perteneciente a Luján. Los resultados obtenidos permitirán obtener, por primera vez, información referente a la composición de la co munidad de hongos liquenizados locales.
MATERIAL Y MÉTODOS
Área de estudio
El área de estudio corresponde al Partido de Luján, ubicado en la Provincia de Buenos Aires al Oeste de la Ciudad de Buenos Aires, entre los 34° 20’ - 34° 54’ Latitud Sur y los 58° 55’ - 59° 20’ Longitud Oeste. Cuenta con una superficie de 777,13 km2 (Bonvecchi et al., 2006). Tiene una población estimada de 120.000 habitantes y se encuentra ubicada a 67 km de la ciudad de Buenos Aires, República Argentina. Este parti do se asienta sobre la Pampa Ondulada (Ferro & Minaverry, 2019) y forma parte de la cuenca media del río Luján. El clima, de acuerdo con la clasificación climática de Köepen, es templa do húmedo con veranos muy cálidos (Carballo & Goldberg, 2014). El partido de Luján forma parte de una gran matriz agropecuaria donde los cultivos son los que ocupan la mayor superficie (Bonvecchi et al., 2006).
Utilizando como base el Código de Ordenamiento Urbano (2019) del partido de Luján, en el presente trabajo se relevaron tres zonas: Casco Fundacional Extendido (CFE), Zona Agropecuaria Extensiva (ZAE), Zona Residencial (ZR). En base a esta Zonificación, la ZAE es definida como promotora del desarrollo de actividades agropecuarias intensivas y de la transición entre la ciudad y el campo. La zona del CFE contiene el casco fundacional de la localidad de Luján e incluye la zona residencial adyacen te que presenta la mayor actividad comercial, de tránsito y actividades humanas de la ciudad de Luján (Fig. 1). Las ZR son aquellas en las que se prioriza la localización de viviendas con el fin de garantizar y preservar las buenas condiciones de habitabilidad. Para este estudio escogimos a Villa Flandria Norte (Localmente denominada “Pueblo Nuevo”) como representante de la ZR, los campos experimentales de la Universidad Nacional de Luján (UNLu) como representante de la ZAE y el Casco Urbano de la localidad de Luján, como representante de la zona del CFE (Fig. 1).
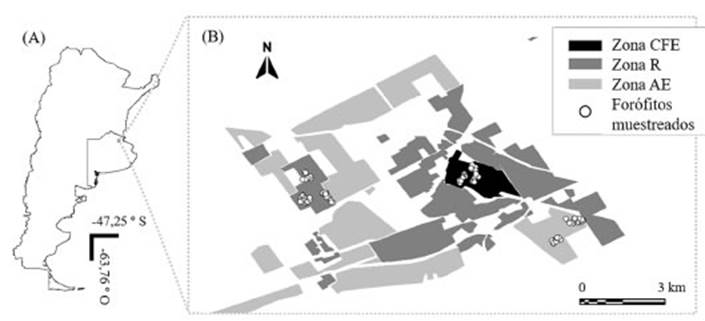
Muestreo
Cada una de las zonas de estudio fue dividi da en celdas de 0,25 km2. Dentro de cada área se seleccionaron al azar tres celdas. Estas celdas fueron denominadas “sitios”. Dentro de cada uno de los sitios se seleccionaron entre 7 y 20 forófitos (árboles en los que crecen los hongos liquenizados) (Fig. 1) que cumplieron con los siguientes requisitos: organismo aparentemen te vivo, diámetro del tronco a altura del pecho (1,50 m) igual o mayor a 10 cm, sin daños en la corteza particularmente en la zona de extracción de muestras, y con tronco sin pinturas ni carte lerías sobre su superficie. El diseño original del muestreo implicaba la selección de 20 árboles por celda. Sin embargo, dada que la elección de las celdas fue al azar, en número de árboles disponi bles fue variable, encontrando en una de las cel das un máximo de 7 árboles que cumplían con to dos requisitos mencionados. La conformación de especies del arbolado en las zonas bajo estudio es significativamente heterogénea y no fue posible utilizar una sola especie arbórea para muestrear en todos los tipos de uso de suelo (ver Análisis estadístico).
Durante el muestreo se determinó la presen cia de cada especie de hongo liquenizado crecien do sobre cada forófito muestreado. Los datos se obtuvieron mediante el uso de un rectángulo de cartón de 10×50 cm (Ochoa-Jiménez et al., 2015). Este rectángulo se colocó sistemáticamente en el lado del tronco con mayor cobertura de líquenes visibles en cada árbol (Sáenz et al., 2007), con el borde superior a 1,5 m sobre el suelo (Sáenz et al., 2007; Estrabou et al., 2005). Se realizaron en total siete campañas a lo largo de seis meses (2019-2020). La información obtenida por rec tángulo fue la siguiente: fecha de registro, ubi cación geográfica, tipo de corteza del forófito (ru goso, liso), forófitos caducifolios o perennes y por último la presencia de cada una de las especies de hongos liquenizados identificados. La iden tificación de los hongos liquenizados se realizó utilizando como referencia la colección deposita da en “Laboratorio B” perteneciente a la UNLu. Además, se efectuaron reacciones puntuales de color con hidróxido de potasio (K) y de hipoclori to de sodio (C) para la identificación de las sus tancias liquénicas las cuales además han sido uti lizadas con mucha frecuencia para la identifica ción de especies (e.g., Suebsri & Wanchai, 2021; Fayyaz et al., 2022; Rosli & Zulkifly, 2022). Los diferentes caracteres morfológicos (Grassi, 1950; Osorio, 1976; Adler, 1992; Scutari, 1992; Scutari, 1995; Scutari & Theinhardt, 2001; Calvelo & Liberatore 2002) se observaron mediante micros copio estereoscópico Zeiss STEMI 2000C. Por úl timo, se construyó una curva de acumulación de especies para determinar si el muestreo fue ca paz de relevar la totalidad o la mayor parte de las especies del conjunto de las zonas de muestreo.
Análisis estadístico
Se aplicó un escalamiento multidimensional no métrico utilizando dos índices de asociación (Bray-Curtis y Morisita). En el nuevo espacio de ordenación se representaron los sitios ordenados según la frecuencia de cada una de las especies de hongos liquenizados registrados (número de foró fitos que presentaron presencia de una especie). Solo se representaron en el espacio de ordenación los vectores de aquellas especies con un aporte sig nificativo a la construcción del espacio de ordena miento, traducido en un valor de p<0,05. En este análisis se incluyeron covariables complemen tarias en cada una de las celdas con el objetivo de determinar si alguna de las mismas presenta algún efecto sobre la ordenación. Las covariables incluidas fueron: (1) frecuencia de forófitos con corteza lisa, (2) frecuencia de forófitos con corte za rugosa, (3) frecuencia de forófitos perennes y (4) frecuencia de forófitos caducifolios. Se realizó un análisis de especies indicadoras medido con el coeficiente phi de Pearson para explorar qué es pecies de hongos liquenizados están relacionadas con cada una de las zonas (Wietrzyk-Pełka et al., 2018). Los análisis estadísticos se realizaron con CRAN R-3.4.2.
RESULTADOS
Se obtuvieron un total de 202 registros (ocu rrencias de especies) de hongos liquenizados en el área de estudio, 55 de los cuales correspon dieron al Casco Fundacional Extendido, 75 a la zona Residencial y 72 a la zona Agropecuaria Extensiva (Tabla 1). Un total de 23 géneros fueron registrados. Los géneros más frecuen tes fueron Physcia (Michaux, 1803) con 39 re gistros (CFE: 14, ZR: 15, ZAE: 10) Candelaria (Massalango, 1852) con 26 registros (CFE: 16, ZR: 9, ZAE: 1), Phaeophyscia (Moberg, 1977) con 23 registros (CFE: 16, ZR: 4, ZAE: 3), Caloplaca (Fries, 1860) con 21 registros (CFE: 0, ZR: 12, ZAE: 9) e Hyperphyscia (Argoviensis, 1894) con 16 registros (CFE: 4, ZR: 7, ZAE: 5). En total, se encontraron 41 especies de hongos liquenizados. La curva de acumulación de especies indica que a través del muestreo se han relevado la mayoría de las especies del área de estudio (Fig. 2).
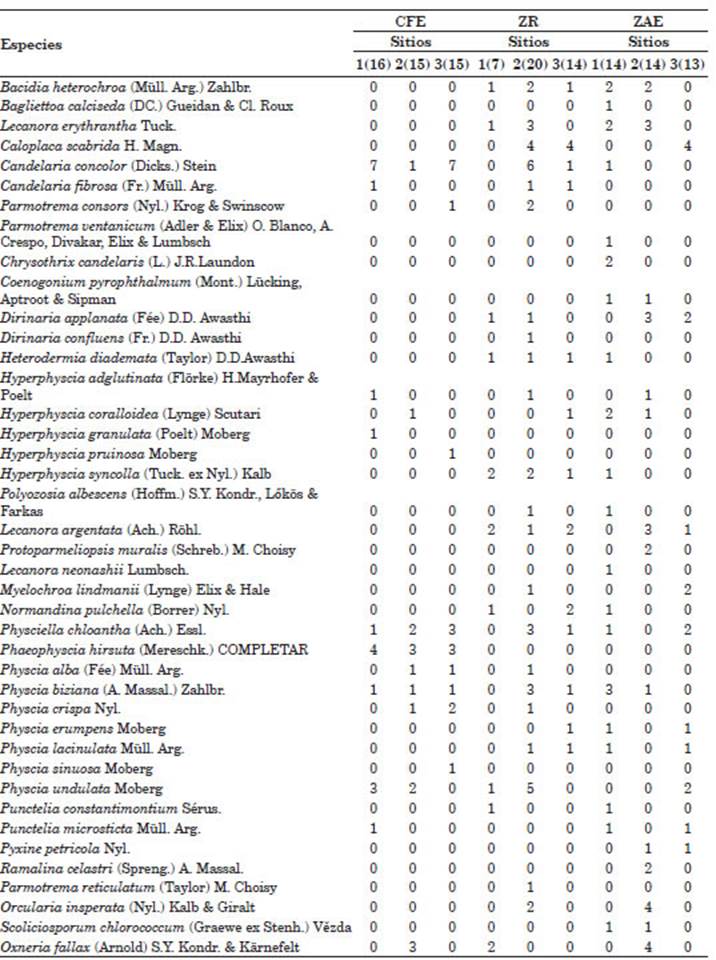
Tabla 1 Especies registradas en el área de estudio. Se presenta la frecuencia (número de árboles o forófitos en los que se ha registrado). Entre paréntesis se indica el número de forófitos muestreados. Abreviaturas: CFE, Casco Fundacional Extendido; ZR, Residencial; ZAE, Agropecuaria Estricta.
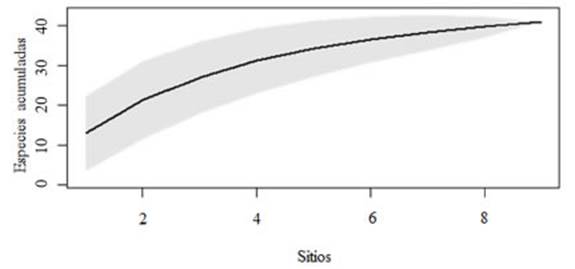
Fig. 2 Curva de acumulación de especies para los 9 sitios relevados en toda el área de estudio (todas las zonas en conjunto).
El análisis de ordenamiento indica que el Casco Fundacional Extendido se separó clara mente de las restantes zonas (Fig. 3). Ninguna de las covariables aporto significativamente al espacio de ordenamiento (p>0,05). Las zonas Residencial y Agropecuaria Extensiva no pu dieron ser claramente separadas en el espacio de ordenamiento (Fig. 3). El Casco Fundacional Extendido y la zona Residencial presentaron solo una especie indicadora, Phaeophyscia hirsuta (Esslinger, 1978) y Hyperphyscia Syncolla (Kalb, 1983)., respectivamente (Tabla 2). El análisis de escalamiento multidimensional no métrico mos tró un resultado similar donde Phaeophyscia hir suta está fuertemente vinculada a los sitios den tro de la zona del Casco Fundacional Extendido. Sin embargo, a través de este análisis, no se ob serva una clara separación entre los sitios dentro de la zona Agropecuaria Extensiva con respecto a la zona Residencial (Fig. 3). Al observar la dis tribución de los sitios y su relación con las espe cies con respecto al primer eje de coordenadas, podemos observar que las especies Caloplaca scabrida (Magnusson, 1950) y Physcia lacinulata (Argoviensis, 1891) están relacionadas negativa mente con dicho eje y se presentan en oposición a los sitios correspondientes al Casco Fundacional (Fig. 3). La especie C. scabrida también se pre senta como especie indicadora de la intersección entre las zonas Agropecuaria y Residencial (Tabla 2). Se registraron tres especies comunes entre la zona Residencial y la Agropecuaria Extensiva (Tabla 2). La zona Residencial comparte una única especie con la zona del Casco Fundacional Extendido: Candelaria concolor (Arnold, 1879); mientras que la zona Agropecuaria Extensiva no presenta especies indicadoras (Tabla 2).
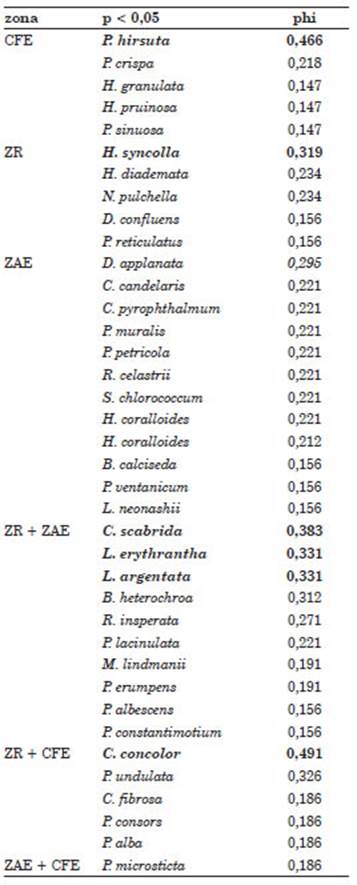
Tabla 2 Resultado del análisis de especies indi cadoras medido con el coeficiente phi de Pearson para explorar que especies de líquenes estaban relacionadas con cada una de las zonas. En neg rita las especies indicadoras para cada zona (o combinación de zonas). Referencias: CFE, Casco Fundacional Extendido; ZR, Residencial; ZAE, Agropecuaria Estricta.
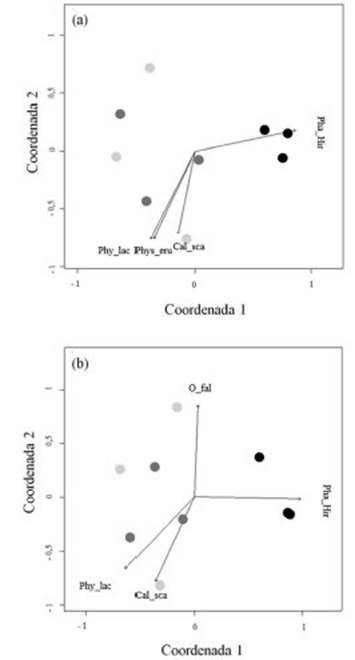
Fig. 3 (A-B) Escalamiento multidimensional no métrico de los sitios de muestreo, ordenamos según la frecuencia de ocurrencia de las especies de líquenes registrados en los forófitos: índices diferentes de similitud (A) Bray-Curtis y (B) Morisita. Abreviaturas: Pha_Hir, Phaeophyscia hirsuta; Cal_sca, Caloplaca scabrida, O_fal, Oxenaria fallax; Phy_lac, Physcia lacinulata; Phys_eru, P. erumpens.
DISCUSIÓN
Casi todas las especies indicadores registra das aquí han sido frecuentemente relacionadas a usos del suelo relacionado a actividades humanas (ver descripción de especies). Este tipo de espe cies pueden tolerar altos niveles de contamina ción, pero desaparecerían si las condiciones cam bian (Fernández-Salegui et al., 2007). Haremos una breve mención a cada una de ellas. Entre pa réntesis, a continuación del nombre de la especie, se indica a que zona está se relaciona la misma.
Phaeophyscia hirsuta (CFE). Esta espe cie es cosmopolita (Infraestructura Mundial de Información en Biodiversidad, 2022). Desde los años 80 se reconoce a P. hirsuta como una es pecie afín a sustratos con enriquecimiento por nutrientes (Atienza & Barreno, 1989; Frati et al., 2007). McCune et al. (1998) determinó que esta especie es indicadora de zonas Urbanas e Industriales en Colorado, Estados Unidos. Munzi et al. (2014) incluyen a P. hirsuta como especie indicadora de un área metropolitana en Lisboa en el suroeste de Europa. Esto es coincidente con los resultados encontrados en el presente tra bajo, donde P. hirsuta es la única especie indica dora del área del Casco Fundacional Extendido (Tab. 2). Según Scutari (1995), esta especie es rara en la provincia de Buenos Aires, creciendo mezclada con los talos de P. chloantha. En la lo calidad de Luján, esta especie solo ha sido regis trada en el Casco Fundacional Extendido (Tab. 1). En la República Argentina, P. hirsuta, ha sido registrada en ambientes relativamente poco per turbados como Sierra Chica, Córdoba, Argentina (Estrabou & García, 1995), así como como en am bientes urbanos (García, 2018).
Hyperphyscia syncolla (ZR). Esta especie ha sido mencionada en otros estudios como indica dora de zonas urbanas (Estrabou et al., 2005; Canseco et al., 2006; Filippini, 2017). Nuestros resultados soportan esta afirmación, aunque par cialmente. En nuestro estudio, H. syncolla no fue registrada en el Casco Fundacional Extendido. En cambio, esta especie fue registrada frecuen temente en la zona Residencial (Tabla 1). Según Estrabou et al. (2005) aunque las zonas urbanas pueden reducir la fertilidad de esta especie, esto no interfiere en la presencia de la misma. Este re sultado, se opone a la ausencia de H. syncolla en el Casco Fundacional. Sin embargo, como men ciona Filippini (2017) es posible que existan fac tores desconocidos hasta el momento que puedan reducir o eliminar la presencia de esta especie en algunas zonas urbanas. Esta última autora, por ejemplo, registró la presencia de espacios verdes urbanos como un factor que impacta negativa mente en la presencia de H. syncolla.
Candelaria concolor (ZR + CFE). Esta es una especie cosmopolita (Er et al., 2015) fre cuente en ambientes urbanos, considerada toxi tolerante, capaz de colonizar en los límites de los desiertos liquénicos (Conti & Cecchetti, 2001; Nimis et al., 2002; LijteRoff et al., 2009). Su frecuencia disminuye al aumentar la ruralidad de las zonas de estudio, es decir, desde la zona del Casco Fundacional Extendido hasta la zona Agropecuaria Extensiva, pasando por la zona Residencial (Tabla 1). Esto coincide con los de scripto por Filippini (2017), donde la frecuencia de C. concolor disminuye al aumentar las áreas cultivadas. Una posible razón de este patrón puede estar relacionada con el aumento del pH en los sustratos dentro de las matrices agro pecuarias, donde la producción de compuestos nitrogenados, como el amoníaco, es dominante (Sparrius, 2007). Candelaria concolor es una es pecie que frecuentemente se asocia a sustratos ácidos (Reed, 2006) por lo que su presencia de bería verse afectada en matrices agropecuarias donde el pH de los sustratos aumenta.
Lecanora argentata (Röhling, 1813) (ZR + AE). Esta especie cosmopolita de distribución templada a subtropical ha sido registrada fre cuentemente en forófitos con corteza lisa, ligera mente ácida, caducifolios, en ambientes abiertos y bien iluminados (Malíček, 2014). En la Argentina ha sido encontrada en regiones templadas a subtropicales de las provincias fitogeográficas de las Yungas, Puneña, Chaqueña y Pampeana (Lumbsch, 1994; Guderley & Lumbsch, 1999; De la Rosa & Messuti, 2014). Habita en sustra tos moderadamente enriquecidos con nutrientes (Ungethüm et al., 2011). Esta especie se propaga en hábitats poco a moderadamente perturbados (Agnan et al., 2017).
Caloplaca scabrida (ZR + ZAE).Osorio (1970) reportó a esta especie por primera vez en la provincia de Buenos Aires sobre corteza. Desde entonces, esta especie solo ha sido regis trada en las provincias de Entre Ríos y Buenos Aires (Liberatore et al., 2012). También ha sido registrada en Uruguay (Osorio, 1970). Se cree que es una especie toxitolerante, debido a que soporta grados de contaminación que son letales para otras especies (Pérez de la Torre, 1985).
Lecanora erythrantha (Tuckerman, 1862) (ZR + ZAE). Esta especie habita en zonas tropicales, subtropicales y templadas de América (Wetmore, 2007). En la Argentina ha sido fre cuentemente registrada en Buenos Aires (Rosato, 2006; Rosato & García, 2014). Hasta donde es conocimiento de los autores, no existe informa ción referente a su toxitolerancia.
Las especies Oxneria fallax (Kondratyuk & Kärnefelt, 2003) y Physcia lacinulata no fueron escogidas como especies indicadoras. Sin embar go, aportan significativamente al ordenamiento generado cuando se utilizó el índice de similitud Morisita. Oxneria fallax se asocia fuertemen te con uno de los sitios agropecuarios (Fig. 3). Esta especie es caracterizada como nitrófila y frecuente en ambiente agropecuarios (LeBlanc & Sloover, 1970; Kondratyuk, 1997; Thor, 1998). No se ha encontrado información referente a la toxitolerancia de P. lacinulata. En el mismo aná lisis de ordenamiento esta especie se encontró fuertemente vinculada a C. scabrida (Fig. 3b). La posición en el espacio de ordenamiento de P. lacinulata y C. scabrida, sugiere que estas dos especies ocupan nichos similares, por lo que po dríamos inferir que P. lacinulata es una especie toxitolerante. A pesar de esta inferencia, será necesario realizar más estudios al respecto para tener una respuesta concreta.
No se pudo identificar alguna especie indica dora de la zona Agropecuaria Extensiva. Este pa trón podría tener su origen en la identidad de las zonas de estudio. Si observamos los resultados de las especies indicadoras (Tab. 2) y el análisis multidimensional no métrico (Fig. 3), es clara la dificultad para separar la zona Residencial de la Agropecuaria Extensiva. La identidad de los sitios está determinada por (1) la caracterización dada por el código de ordenamiento urbano, es decir, si es considerada Casco Urbano, Residencial o Agropecuario por motivos catastrales y (2) la in fluencia ambiental que tiene cada tipo de uso del suelo sobre los forófitos relevados. Sin embargo, cuando un forófito relevado se encuentra cerca o en el límite de zonas según lo especifica el código de ordenamiento urbano, resulta que la influen cia del uso del suelo sobre el forófito no tiene un solo origen. En consecuencia, aquellos forófitos que se encuentran, por ejemplo, cerca del límite de la zona Residencial, pueden presentar mayor influencia de la zona Agropecuaria que de la mis ma zona Residencial. Esto parece ser muy común en la localidad de Luján, donde los ejidos urba nos se desarrollan dentro de una matriz agrope cuaria extensa y el paso de zonas residenciales a agropecuarias es, en muchos casos, abrupto. Esto apoya los resultados que muestran la difi cultad para diferenciar las zonas Residencial de la Agropecuaria Extensiva según su composición liquénica. La influencia de la actividad agrope cuaria en la zona Residencial de muestreo parece ser lo suficientemente grande como para que no se la pueda distinguir de la zona Agropecuaria Estricta.
CONCLUSIONES
En el presente trabajo se logró determinar cuáles son las especies asociadas a diferentes usos del suelo en el partido de Luján. Además, se presenta por primera vez una lista de especies de hongos liquenizados cortícolas de dicho partido, ampliando la información preexistente. En base a estos resultados consideramos que este trabajo puede aportar al conocimiento de la distribución de las especies de hongos liquenizados de la pro vincia de Buenos Aires (Argentina). Respecto a las especies indicadoras, con alguna excepción, la mayoría de estas especies son consideradas toxi tolerantes en diferente grado. Este resultado, aunque nuevo, no es sorprendente. El partido de Luján se caracteriza por su predominante activi dad agropecuaria. Muchas localidades dentro del partido son relativamente pequeñas comparadas con la superficie utilizada para actividades agro pecuarias. Las especies de hongos liquenizados cortícolas en el partido se enfrentan, por un lado, a la contaminación asociada a las zonas urbani zadas intensamente, como es el caso del Casco Urbano de la localidad de Luján y, por otro lado, al enriquecimiento por nutrientes de las cortezas de los árboles en las zonas de uso agropecuario en los que se desarrollan. Estas condiciones am bientales influyen en la composición de las es pecies locales dejando lugar solo a especies que se encuentran relacionadas fuertemente con las actividades humanas.

