











 texto en
texto en  Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
A pesar de los múltiples avances en los diver sos campos de la biología humana y cuando pareciera que, hoy, al hombre es posible representarlo con una traza infinitesimal de su genoma, es curioso, sin embar go, que aún haya regiones de la anatomía humana que sean motivo de controversias, desacuerdos y malen tendidos. Esto es lo que ocurre, desde hace décadas, si no siglos, con la anatomía de la unión gastroesofágica, con los elementos que la componen y de aquellos que la circundan. Por lo que resulta que, si la anatomía es ignorada o mal comprendida, resulta más difícil poder entender cómo es que todo funciona normalmente o cuándo enfrentamos su disfunción y enfermedad y, por último, las consecuencias que, para bien o para mal, tienen nuestras intervenciones sobre ella.
En este artículo revisaremos algunos aspectos embriológicos, anatómicos y fisiológicos que nos pare cen relevantes y revisten interés para el cirujano que aborda esta región.
El diafragma
El diafragma es una delgada lámina musculo tendinosa que separa la cavidad torácica de la abdo minal. Con una forma elíptica constituye la separación entre el tórax y el abdomen; tiene en su centro un área tendinosa con dos cúpulas de músculo estriado que se alzan a cada lado. Se inserta por adelante en el xifoides, hacia lateral en la superficie interna de los cartílagos costales de la séptima a la duodécima costilla y por atrás en las apófisis transversas de la primera vértebra lumbar y en el cuerpo y sínfisis de las tres primeras lumbares.
Embriológicamente, el diafragma se origina de cuatro estructuras: el septum transverso, las membra nas pleuroperitoneales, el mesenterio dorsal del esófa go y los músculos de la pared. A la cuarta semana de desarrollo, el septum transverso de origen mesodérmi co separa de forma incompleta las cavidades pericárdi cas de las peritoneales e inicia su migración hacia distal siguiendo el crecimiento del embrión y arrastrando los nervios frénicos. A la octava semana, el septum alcanza su ubicación toracolumbar, pero no cierra toda el área pues la comunicación permanece a través de los lla mados ductos pleuroperitoneales; en ese período las membranas pleuroperitoneales derecha e izquierda se fusionan al septum y al mesenterio dorsal del esófago completando la separación de las cavidades pleurales de la abdominal. El septum dará origen al tendón cen tral y a los músculos más centrales. Los músculos poste rolaterales provienen de la pared. El mesenterio dorsal del esófago aportará las fibras musculares que desarro llarán el área de la crura. Este origen diverso determina el llamado diafragma costal y el crural.
Su irrigación depende de va sos pericardiofrénicos, mamaria inter na y ramos de aorta torácica y abdominal.
La inervación motora del diafragma proviene únicamente del nervio frénico homolateral y de esta inervación también depende la región crural. Es impor tante conocer la distribución de la inervación frénica para no lesionarla cuando se secciona el diafragma. Es posible la sección en la periferia del diafragma cerca na a la inserción costal y también algunos centímetros en el borde anterior del hiato esofágico hacia el centro frénico para avanzar en el mediastino en la llamada ma niobra de Pinotti1-4.
El hiato
El diafragma es atravesado por varias estruc turas, pero tres son las principales y tienen aperturas propias: la aorta, la vena cava y el esófago. La aorta lo hace a través de un orificio localizado a nivel de T12 o L1, limitado hacia dorsal por los cuerpos vertebrales, la teralmente por los pilares o cruras diafragmáticas y por anterior por el ligamento arcuato medio que une los dos pilares. Junto con la aorta atraviesa el conducto to rácico y ocasionalmente las venas ácigos y hemiácigos.
El foramen de la vena cava está a nivel de T8 y se ubica a la derecha de la línea media en pleno tendón central, por lo que todos sus márgenes son tendinosos. El nervio frénico derecho y algunos linfáticos la acompañan.
El hiato esofágico se ubica a nivel de T10, ligera mente a la izquierda y ventral a la aorta. A diferencia de las anteriores, esta apertura es la única completamen te formada por musculatura. De acuerdo con algunos autores existen múltiples variantes en la conformación muscular del hiato pero, en más del 60%, la crura dere cha se divide y rodea por completo el hiato y se suma a la crura izquierda, por lo que esta suele verse más firme y gruesa que la derecha (Fig. 1). Por el hiato cruzan los nervios vagos, algunos ramos arteriales y venosos gás tricos izquierdos y del nervio frénico izquierdo.
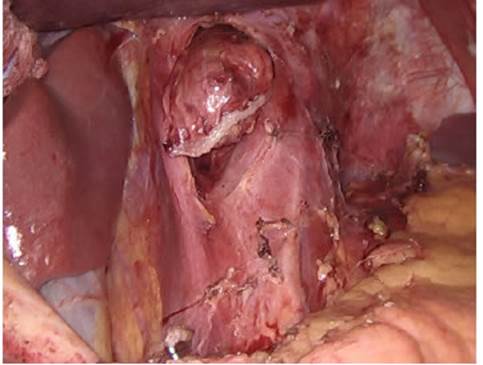
La porción crural del diafragma cumple un papel relativamente menor en la función respiratoria comparada con la porción costal, pero sí desempeña un importante papel en la función gastroesofágica, parti cularmente en la deglución, el vómito y en la preven ción del reflujo gastroesofágico.
La región crural está coordinada con el mo mento de la deglución, de modo tal que la crura se rela ja transitoriamente cuando el bolo alimenticio distien de el esófago y es propulsado por la peristalsis a fin de facilitar y permitir su tránsito a través del hiato.
Durante la emesis se requiere una compleja coordinación de la musculatura respiratoria y abdomi nal, del diafragma y la esfagogástrica. En la fase expul siva, la porción costal del diafragma se mantiene con traída a fin de generar presión positiva intrabdominal, mientras que la porción crural se relaja para permitir la salida del contenido gástrico1,2. Hay autores que consi deran la crura diafragmática de gran importancia en la prevención del reflujo gastroesofágico y hay quienes la consideran como un segundo esfínter5. Parece razona ble que, por su estructura muscular y ubicación respec to de la unión gastroesofágica, la crura actúe como un apoyo externo previniendo el reflujo en condiciones de aumento de presión abdominal; sin embargo, a nues tro entender, no parecería tener un papel antirreflujo determinante, en condiciones normales de reposo o interdigestivo.
La membrana frenoesofágica
Se denomina esófago abdominal el segmento de 2 a 4 cm de tubo esofágico, que es posible exponer y rodear, situado entre el hiato diafragmático y su unión con el estómago. El esófago se identifica por carecer de serosa y las fibras longitudinales de su capa exter na. Su unión con el estómago se denomina cardias y se reconoce como el límite oblicuo entre la musculatura esofágica y la serosa gástrica que se extiende desde el ángulo de His hacia la curva menor, proximal a la vena cardiotuberositaria.
El hiato diafragmático, el esófago abdominal y el cardias están ocultos por hallarse cubiertos por un manguito formado por una membrana fibroelástica denominada ligamento o membrana frenoesofágica. Además, en el abdomen, esta estructura se encuentra recubierta por el peritoneo (Fig. 2).
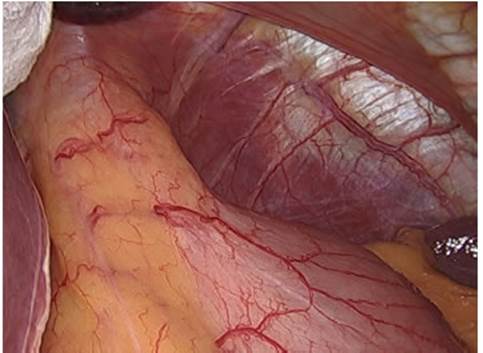
Figura 2 Membrana frenoesofágica recubierta por peritoneo, con grasa en su espesor (fat pad), que mantiene oculta la unión gastroesofágica.
La membrana frenoesofágica está compuesta de fibroblastos, colágeno y fibras elásticas y tiene su origen en la prolongación de las fascias endotorácica y tranversalis, las que se fusionan a nivel del hiato; una hoja ascendente se inserta en la pared esofágica 2 a 4 cm proximal al hiato, mientras la hoja descendente lo hace en la serosa del fondo gástrico. Suele tener un co jinete graso en su espesor denominado fat pad, y por ella transitan los nervios vagos (Fig. 3).

Figura 3 Esquema que muestra la estructura de la membrana frenoesofágica, su relación con el hiato y los elementos de la unión gastroesofágica
Constituye un sello que separa el tórax del abdomen a nivel del hiato y además es un anclaje del esófago al hiato, pero que le permite cierta movilidad durante la deglución.
Para remover esta membrana y despejar la unión gastroesofágica se puede abordar desde la curva menor o desde el ángulo de His (esto último parece más recomendable) y desde allí disecarla hacia medial, libe rando el pilar izquierdo y protegiendo el vago anterior.
Hay autores que le atribuyen un papel en el mecanismo antirreflujo y también en su eventual rela ción con las hernias hiatales6,7.
El esfínter esofágico inferior o gastroesofágico
Pocas estructuras en la anatomía humana han sido y son más debatidas que la existencia de un es fínter anatómico a nivel de la unión gastroesofágica (UGE)8, esto a pesar de que los estudios manométri cos han demostrado un mecanismo esfinteriano en la región9,10 y que constituye, por lo demás, la principal barrera contra el reflujo gastroesofágico11. El gran pro blema, hasta años recientes, ha sido demostrar una estructura anatómica en el esófago distal que coincida con el concepto clásico, y arbitrario vale agregar, de esfínter, esto es, un anillo muscular formado por un músculo circular distinguible, ojalá palpable y separa do de los músculos adyacentes por tabiques de tejido conectivo12. Tal estructura nunca se ha demostrado en la UGE, aunque algunos autores a lo largo de la historia han creído o imaginado encontrarla8. La ausencia de tal estructura llevó a sostener la inexistencia de un esfínter anatómico a ese nivel, pero debido al trabajo de Fike y col. en 19569, que confirmaba la existencia de un esfín ter manométricamente demostrable a nivel de la UGE, llevó a los especialistas a plantear la insólita e inédita interpretación, prevalente aún en nuestros días, de la existencia de un esfínter fisiológico pero no anatómi co. Winans, en 1977, desafió todavía más el concepto clásico del esfínter circular, y de paso el “fisiológico”, cuando demostró la existencia de una asimetría mano métrica del esfínter esofágico inferior13.
En 1979, los estudios anatómicos de la UGE de Liebermann y col.14 zanjaron el asunto al demostrar que el esfínter esofágico inferior no está formado por un músculo anular sino por dos bandas musculares que cierran el lumen actuando complementariamente: las fibras “clasp” por curvatura menor y las fibras “sling” u oblicuas por curva mayor. La descripción señala una particular disposición y un incremento en la cantidad de fibras de la capa muscular interna en la UGE. Las fi bras de la capa muscular interna no forman un anillo o músculo circular que rodee todo el perímetro de la región del cardias; en cambio se observa un área de fibras semicirculares o corchetes orientados transver salmente que abrazan la curvatura menor en la UGE; estas fibras llamadas “clasp” se insertan firmemente en el tejido conectivo submucoso en el margen de contac to de las fibras oblicuas. Las fibras “sling” u oblicuas se despliegan por el lado de la curvatura mayor (Fig. 4), constituyen un haz muscular de 3 cm de ancho y cubren un área que comienza 1,5 cm proximal al ángulo de His formando parte del extremo distal del esófago, rodeando la UGE a manera de una estola; sus dos brazos ‒anterior y posterior‒ corren paralelos a la curvatura menor terminando en el antro. El extremo de las fibras “clasp” se topan casi en ángulo recto en el margen de las fibras oblicuas.

Figura 4 Disposición de las fibras semicirculares “clasp” por curva menor y las fibras oblicuas “sling” por curvatura mayor. Estas bandas muscula res constituyen la estructura anatómica del esfínter gastroesofágico. Nótese que no existen fibras que rodeen el perímetro de la unión gastroesofágica15
Por lo tanto, el EEI no es un esfínter anular sino está formado por dos haces de músculos que actúan de forma complementaria en el cierre del lumen: las fibras musculares “clasp“ y las “oblicuas o cling”. Varios trabajos de nuestro grupo confirman estos hallazgos y descartan definitivamente la existencia de un músculo anular15.
Comprender la anatomía del esfínter esofágico inferior con su particular estructura muscular, eliminan do el concepto de un esfínter anular o solo fisiológico, es muy importante, porque permite entender por qué las acciones quirúrgicas, no solo sobre el cardias sino incluso sobre el cuerpo gástrico, pueden tener reper cusión en la acción esfinteriana. La ubicación de la car diomiotomía de Zaijer, ahora el POEM, en la acalasia, no es indiferente16, así también la gastrectomía par cial distal o la gastrectomía vertical de la manga pue den incidir y comprometer seriamente la competencia esfinteriana17.

